Voltar
ESTRUTURA
DA VEGETAÇÃO E CONDIÇÕES EDÁFICAS NUMA CLAREIRA DE MATA
DE RESTINGA NA RESERVA BIOLOGICA ESTADUAL DA PRAIA DO SUL
(RJ)
Dorothy Sue
Dunn de Araujo1, Rogério Ribeiro de Oliveira1,
Eduardo Lima2, Alexandre Ravelli Neto2
1Serviço de
Ecologia Aplicada, Divisão de Estudos Ambientais, Fundação
Estadual de Engenharia do Meio Ambiente (FEEMA), Estr. Vista
Chinesa, 741, Rio de Janeiro, RJ, CEP 20531-410.
2Instituto
de Agronomia, Departamento de Solos, Universidade Federal
Rural do Rio de Janeiro, Estr. Velha Rio-São Paulo, Km 47,
Seropédica, RJ. CEP 23851-970.
RESUMO
Na planície arenosa
da Praia do Sul (Ilha Grande - RJ), várias estradas foram
abertas em 1980, e posteriormente abandonadas. Oito anos
depois da derrubada da mata foram amostrados os indivíduos
com DAP ³ 2,5cm e altura ³ 2m num total de 30 parcelas (5
x 5m) localizadas ao longo da estrada principal. Na área
em regeneração, das 18 espécies ocorrentes a mais importante
foi Byrsonima sericea DC. seguido por Cecropia
lyratiloba Miq. e C. glazioui Snethl. A serapilheira
acumulada no solo da área em regeneração foi de 509 ± 238g.m-2
versus 920 ± 170g.m-2 na mata original. À exceção do elemento
fósforo, todos os nutrientes do solo apresentaram valores
mais elevados na mata. As variações de temperatura do solo
nos dois ambientes foram muito mais acentuadas na área em
regeneração. A capacidade de retenção de cátions do solo
sob mata foi 2 vezes superior quando comparado ao solo da
área em regeneração. Uma possível explicação para o menor
desenvolvimento da vegetação é o baixo teor de matéria orgânica
(1,15%) verificada nesta área. Passados 8 anos da abertura
da estrada, com as ações de distúrbio coibidas, e ainda,
com a grande probabilidade de disseminação de propágulos
da mata, foi demonstrado que as restingas constituem ecossistemas
de regeneração muito lenta.
Palavras-chave:
mata de restinga, distúrbio, fatores edáficos, reserva biológica.
INTRODUÇÃO
Uma parte significativa
das árvores de matas tropicais úmidas requer a existência
de uma abertura no dossel para que elas possam se regenerar
(11, 15). Estas aberturas modificam as características do
meio, propiciando um aumento dos níveis de luminosidade,
temperatura do ar e do solo, entrada de chuva e disponibilidade
de nutrientes e, ao mesmo tempo, uma diminuição na umidade
relativa do ar (4), que, por sua vez, estimulam as taxas
de substituição natural dos indivíduos (8).
A influência de clareiras
na dinâmica de florestas tropicais é reconhecida pelo menos
desde o estudo pioneiro de Richards (22). Nos últimos 20
anos, um número crescente de estudos tem sido desenvolvido
sobre o assunto no Neotrópico, especialmente na América
Central. Uma extensa revisão do assunto é apresentada (11).
Em contraponto ao processo
natural de formação de clareiras, geralmente causado por
queda de árvores inteiras ou galhos de árvores, existem
as aberturas não naturais, causadas por desmatamentos, incêndios
e/ou trabalhos de terraplanagem, que apresentam uma magnitude
de impacto mais intensa que aqueles de causas naturais.
Este tipo de distúrbio geralmente acarreta a destruição
de todos os indivíduos, bem como do banco de sementes do
solo, além da exposição do solo (no caso de terraplanagem).
Os estudos sobre este tipo de distúrbio têm sido menos freqüentes,
provavelmente devido às dificuldades de manter o controle
destas áreas, uma vez que seu destino é geralmente a utilização
pelo homem.
No entanto, o estudo
de distúrbios antrópicos em formações tropicais é de grande
importância para obtenção de conhecimentos sobre a dinâmica
de regeneração. Estes conhecimentos podem ser aplicados
no manejo de áreas a serem recuperadas e, também, no planejamento
do uso sustentado da floresta tropical. A aplicação das
teorias sobre clareiras ao manejo de florestas tropicais
por meio do conhecimento dos processos de regeneração em
áreas devastadas artificialmente foi feita na Amazônia Peruana
(15, 16), em áreas de floresta sobre solos arenosos e pobres
em nutrientes.
As florestas que ocupam
as planícies arenosas da costa brasileira (matas de restinga)
também são consideradas florestas tropicais úmidas (23).
Entretanto, apresentam um conjunto de processos abióticos
bastante distinto das demais formações de mata tropical
úmida (13, 17).
Com relação a distúrbios
em restingas, autores (3) estudaram a regeneração de uma
comunidade baixo-arbustiva na restinga de Jacarepaguá após
incêndio e Sá (24) descreveu a vegetação seis anos após
sua derrubada por trator em arruamentos feitos na mata de
restinga da Reserva Ecológica Estadual de Jacarepiá, Saquarema
(RJ).
Os objetivos deste trabalho
são a descrição da vegetação e alguns aspectos ecológicos
e edáficos ocorridos no leito de uma estrada aberta na restinga
da Reserva Biológica Estadual da Praia do Sul oito anos
após o distúrbio, comparando-os com a mata intacta contígua.
MATERIAIS E MÉTODOS
A área de estudo está
localizada na Reserva Biológica Estadual da Praia do Sul
(RBEPS), situada na vertente meridional da Ilha Grande,
município de Angra dos Reis, Estado do Rio de Janeiro (23o
10'S; 44o 17'W) (Figura 1). A estação meteorológica mais
próxima encontra-se no continente, na cidade de Angra dos
Reis, a 20km em linha reta do local de estudo. A temperatura
média anual desta é 22,6oC e a precipitação média anual
é de 2.314mm (2). Nos meses de junho, julho e agosto ocorre
uma redução da pluviosidade, não se verificando no entanto
situações de déficit hídrico. Na área da Reserva, a média
de altura pluviométrica entre os anos de 1993 e 1995 foi
de 1.515mm (19). Os ventos são predominantemente dos quadrantes
leste e sul.
A RBEPS ocupa uma área
de 3.600ha, sendo que cerca de 800ha constituem uma planície
com formação atribuída ao Pleistoceno Superior e Holoceno,
a partir de mudanças paleoambientais, principalmente climáticas
e de variação do nível do mar (1). Pode-se identificar quatro
comunidades vegetais nesta planície: psamófila reptante
de anteduna, mata de cordão arenoso, mata alagadiça de planície
e manguezal (2).
A mata de cordão arenoso
é uma floresta de aproximadamente 10m de altura. As copas
das árvores são contíguas na sua maioria, permitindo porém
a passagem de luz suficiente para o estabelecimento de um
estrato inferior constituído por gravatás e samambaias.
As árvores mais comumente encontradas são Rheedia brasiliensis
(Mart.) Planch. & Triana, Tapirira
guianensis Aubl., Ilex integerrima
(Vell.) Reissek e Melanopsidium nigrum Cels. No extrato
arbustivo encontram-se Psychotria carthaginensis
Jacq., P. barbiflora DC., Coccoloba
confusa How., Psidium cattleyanum
Sabine e Gomidesia fenzliana Berg, entre outras (2).
Figura 1.
Mapa de localização da Reserva Biológica Estadual da Praia
do Sul, com indicação da área de estudo.
Em 1980, a planície
da Reserva foi palco de tentativa de um projeto imobiliário
que pretendia lotear toda a área da Baixada da Praia do
Sul. Foram abertas, por meio de uma máquina de terraplanagem,
cerca de 6km de estradas com largura de 5-7m, nas quais
foram removidas a cobertura vegetal e as camadas superiores
do solo. No cordão externo de restinga, foi aberta uma estrada
posicionada paralelamente à linha da praia, a cerca de 100m
desta. Com a criação, em 1981, da Reserva Biológica Estadual
da Praia do Sul, as ações de desmatamento foram imediatamente
sustadas e a área mantida sob estrito controle. Desde então,
as comunidades vegetais sobre o leito das estradas vêm sofrendo
um processo de regeneração natural, cuja estrutura e condições
edáficas são objetos da presente contribuição.
A amostragem da vegetação
foi realizada em 30 parcelas, medindo 5x5m (totalizando
750m2) distribuídas aleatoriamente no leito da estrada abandonada
no cordão externo de restinga. Nestas parcelas foram amostrados
todos os indivíduos com DAP ³ 2,5cm e altura ³ 2m. Para
os indivíduos com mais de um tronco, foi calculado um único
diâmetro baseado nas áreas basais dos vários troncos. Dentro
de cada parcela, foram delimitadas aleatoriamente duas subparcelas
de 1x1m onde foram estimadas visualmente
a cobertura percentual de vegetação herbácea, projeção vertical
da copa (5) e as percentagens do substrato coberto por serapilheira
e de areia exposta.
Na mata de restinga
intacta, em faixa paralela à estrada, foi mensurado o perímetro
de todos os indivíduos com DAP ³ 2,5cm e altura ³ 2m em
um transect de 187,5 x 4m (750m2). Nesta amostragem
foram identificados
apenas os indivíduos de Byrsonimia sericea DC. A
estimativa do número total de espécies que ocorre nesta
mata foi baseada no material botânico identificado a nível
de espécie, que vem sendo coletado na área desde 1983. No
sentido de aproximar as metodologias empregadas, foram considerados
apenas os grupos e formas vegetais amostradas no levantamento
da vegetação da área em regeneração. Os parâmetros fitossociológicos
foram calculados segundo Brower & Zar (5).
Para se medir as temperaturas
do solo nas áreas de distúrbio (leito da estrada abandonada)
e de mata intacta nas profundidades de 5 e 10 cm, foi utilizado
termômetro digital marca Aucon. Foram feitas duas seqüências
de medições: uma diariamente, em um período de 12 dias (de
7 a 18 de agosto de 1989) no horário das 14 horas, e uma
segunda, destinada ao acompanhamento da marcha diária da
temperatura, feita de hora em hora no dia 8 de janeiro de
1991.
Para determinação da
massa de serapilheira acumulada, foram estabelecidos aleatoriamente
2 quadrados (30x30cm) em cada uma das 30 parcelas do leito
da estrada abandonada. Para a mata intacta, em função de
maior homogeneidade do manto de serapilheira, foram feitas
25 amostragens com as mesmas dimensões. Em cada quadrado,
a serapilheira (horizonte 0o) foi removida com auxílio de
faca e recolhida em sacos plásticos etiquetados. Em laboratório,
o material foi colocado em estufa a 80oC até atingir peso
constante. Após pesagem, os resultados foram transformados
para gramas de peso seco de serapilheira por metro quadrado
(gps.m-2).
Foram tomadas amostras
compostas de solo nos dois locais de estudo, segundo a metodologia
de Chapman & Pratt (7) até a profundidade de 20cm. Os
elementos analisados foram: pH, Ca++, Mg++, Al+++, H+, P,
K+, Na+ e C org (12). A partir dos valores obtidos, foi
calculado o valor S (soma das bases) e o valor T (capacidade
de troca de cátions). As análises foram realizadas no Departamento
de Solos da Universidade Federal Rural do Rio de Janeiro.
RESULTADOS E DISCUSSÃO
A vegetação da área
em regeneração é de baixa estatura (altura média = 4,3m)
e apresenta uma densidade total de 1.400 indivíduos vivos
por hectare e uma área basal total de 3,51m2.ha-1 (Tabela
1). De um modo geral, é distribuída de maneira descontínua,
com 57% das parcelas apresentando no máximo dois indivíduos
e o restante tendo uma média de 6,7 indivíduos e englobando
75% da área basal total. O estrato herbáceo é esparso (cobertura
média = 21,2%), porém 75,3% do substrato é coberto por serapilheira.
A presença da mata intacta beirando os dois lados da área
amostrada certamente contribui de maneira significativa
para o acúmulo de detritos no chão. A mata intacta apresenta
uma densidade total de 2.560 indivíduos vivos por hectare,
com uma área basal total de 25,9m2.ha-1
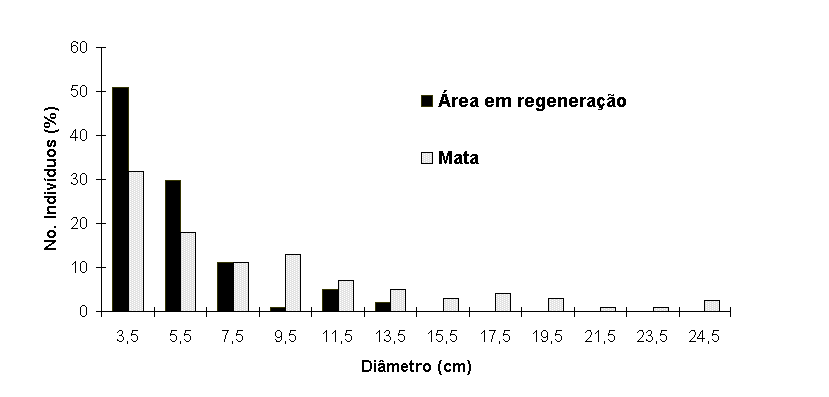
(Tabela 1). É constituída de troncos mais grossos (Figura
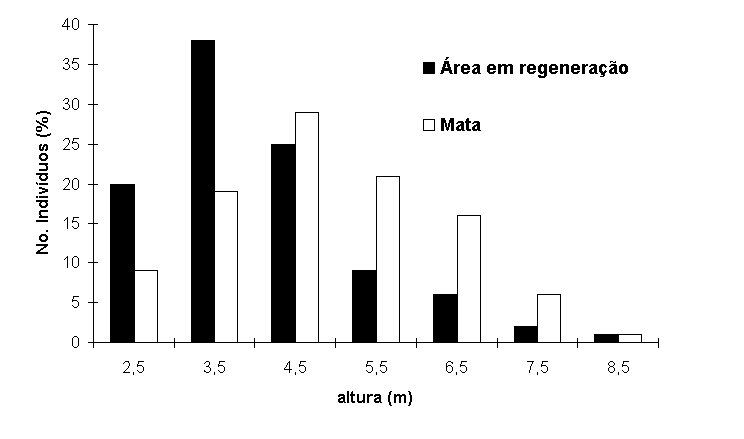
2) e mais altos (Figura 3) que a vegetação da área em regeneração.
TABELA 1: Comparação
entre parámetros da mata intacta e da área em regeneração
na planície arenosa da RBEPS, Ilha Grande, RJ.
| |
Área
em regeneração |
Mata
intacta |
| Indivíduos
vivos |
|
|
| Densidade
(ind./ha) |
1400 |
2560 |
| Dominância
(m2/ha) |
3.51 |
25.9 |
| No.
Indivíduos c/ troncos multiplos (%) |
25.7 |
29.7 |
| Altura
média (m) |
4.3 |
5.2 |
|
|
|
| Indivíduos
mortos |
|
|
| Densidade
(ind./ha) |
26.7 |
200 |
| Dominância
(m2/ha) |
0.04 |
0.87 |
|
|
|
| Byrsonima
sericea |
|
|
| Densidade
(ind/ha) |
680 |
67 |
| Dominância
(m2/ha) |
2.1 |
2.23 |
| No.
Indivíduos c/ troncos multiplos (%) |
45 |
40 |
| No.
espécies |
18 |
64* |
* Estimativa do no.
total de espécies lenhosas encontrado na mata baseada em
coletas aleatórias realizadas durante um período de 10 anos
A espécie dominante
na área em regeneração é Byrsonima sericea DC., com
45% do VI total (Tabela 2). Parece tratar-se de uma espécie
importante no processo regenerativo em áreas de restinga,
pois também foi apontada por (3) como uma das espécies dominantes
30 meses após uma queimada, numa área que sofria periodicamente
a ação do fogo. Entretanto, em outra área de restinga em
regeneração no Município de Saquarema, esta espécie desempenhou
um papel muito menos significativo, com baixa densidade
(85ind/ha) e dominância (1,2m2/ha), 6 anos após
o distúrbio (24).
Figura 2. Histograma
dos diâmetros (DAP ³ 2,5cm) dos indivíduos arbóreos na mata
e na área em regeneração na planície arenosa da Reserva
Biológica Estadual da Praia do Sul, Ilha Grande, RJ.
Figura 3.
Histograma das alturas dos indivíduos arbóreos na mata e
na área em regeneração na planície arenosa da Reserva Biológica
Estadual da Praia do Sul, Ilha Grande, RJ.
Neste estudo, Trema
micrantha (L.) Blume e Aegiphila sellowiana Cham.,
que possuem baixa tolerância à sombra (6) englobaram 22,7%
do VI total. O sucesso destas espécies na regeneração da
mata de restinga em Saquarema pode ser atribuído à maior
largura (20m) e orientação dos arruamentos feitos por trator.
Enquanto na RBEPS a estrada está orientada na direção E-W,
que recebe menos luminosidade (21), a maior parte dos arruamentos
amostrados (62,5%) no estudo referido (24) estão orientados
na direção N-S.
TABELA 2: Parâmetros
fitossociológicos da área em regeneração na planície arenosa
da Reserva Biológica Estadual da Praia do Sul,RJ. Ni = no.
indivíduos; Pi = no. de parcelas com espécie i; FAi = freqüência
absoluta; FRi = freqüência relativa; DAi = densidade absoluta;
DRi = densidade relativa; DoAi = dominância absoluta; DoRi
= dominância relativa; VI = valor de importância.
| Espécie |
Ni |
Pi |
FAi |
FRi |
DAi |
DRi |
DoAi |
DoRi |
VI |
|
Byrsonima sericea
DC. |
51 |
16 |
0,53 |
29,09 |
680,00 |
48,57 |
2,08 |
59,39 |
137,05 |
|
Cecropia lyratiloba
Miq. |
14 |
9 |
0,30 |
16,36 |
186,67 |
13,33 |
0,55 |
15,73 |
45,43 |
|
Cecropia glazioui
Snethl. |
5 |
4 |
0,13 |
7,27 |
66,67 |
4,76 |
0,32 |
9,19 |
21,19 |
|
Amaioua intermedia
Mart. ex R. & S. |
8 |
2 |
0,07 |
3,64 |
106,67 |
7,62 |
0,11 |
3,04 |
14,29 |
|
Ilex integerrima
(Vell.)Reissek |
4 |
4 |
0,13 |
7,27 |
53,33 |
3,81 |
0,04 |
1,18 |
12,26 |
|
Ficus
sp. |
4 |
3 |
0,10 |
5,45 |
53,33 |
3,81 |
0,03 |
0,95 |
10,21 |
|
Solanum caavurana
Vell. |
3 |
3 |
0,10 |
5,45 |
40,00 |
2,86 |
0,04 |
1,06 |
9,38 |
|
Solanum swartzianum
R. & S. |
2 |
2 |
0,07 |
3,64 |
26,67 |
1,90 |
0,08 |
2,28 |
7,82 |
|
Annona
cf. acutiflora Mart. |
2 |
2 |
0,07 |
3,64 |
26,67 |
1,90 |
0,06 |
1,60 |
7,14 |
|
Tabebuia
sp. |
3 |
1 |
0,03 |
1,82 |
40,00 |
2,86 |
0,08 |
2,28 |
6,95 |
|
Tapirira guianensis
Aubl. |
2 |
2 |
0,07 |
3,64 |
26,67 |
1,90 |
0,03 |
0,72 |
6,26 |
| Indeterminada |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,03 |
0,76 |
3,53 |
|
Aureliana
cf. fasciculata (Vell.)Sendtn. |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,02 |
0,46 |
3,23 |
|
Guapira opposita
(Vell.) Reitz |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,02 |
0,46 |
3,23 |
|
Andira fraxinifolia
Benth. |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,01 |
0,30 |
3,07 |
|
Rapanea umbellata
(Mart.)Mez |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,01 |
0,27 |
3,04 |
|
Schinus terebinthifolius
Raddi |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,01 |
0,19 |
2,96 |
|
Ouratea cuspidata
(St.-Hil.) Engl. |
1 |
1 |
0,03 |
1,82 |
13,33 |
0,95 |
0,01 |
0,19 |
2,96 |
| |
105 |
|
|
|
1400 |
|
3,51 |
|
300 |
Na RBEPS, Byrsonima
sericea DC. apresenta uma densidade de apenas 67ind.ha-1
na mata intacta e com área basal um pouco acima daquela
na área em regeneração (Tabela 1). A baixa densidade desta
espécie associada a diâmetros grossos e à falta de indivíduos
jovens em matas de restinga já foi observada na Reserva
Ecológica de Jacarepiá, Município de Saquarema (Sá &
Araujo, dados não publicados), indicando ser uma espécie
que não se reproduz dentro da mata fechada. Ou seja, é uma
espécie pioneira de vida longa no sentido de Whitmore (26),
que atinge o dossel.
É comum em áreas de
restinga em regeneração a ocorrência de indivíduos com vários
troncos rebrotando ao nível do solo ou um pouco acima (24).
Na RBEPS, 25,7% de todos os indivíduos na área em regeneração
apresentaram troncos múltiplos; na mata contígua esta porcentagem
é semelhante (29,9%). Da espécie dominante, B. sericea
DC., 45% dos troncos eram múltiplos.
Das 17 espécies (excluindo
a indeterminada) encontradas no leito da estrada, apenas
6 (35%) são encontradas exclusivamente em ambientes secundários
iniciais na RBEPS (Aureliana cf. fasciculata (Vell.)
Sendtn., Cecropia lyratiloba Miq., C. glazioui
Snethl., Schinus terebinthifolius Raddi, Solanum
caavurana Vell., S. swartzianum Roem. & Schult.);
as outras ocorrem também na mata intacta. É surpreendente
esta baixa diversidade de espécies quando comparada com
outras áreas de mata de restinga em regeneração, como em
Saquarema (24), onde foram encontradas em torno de 50 espécies
em área amostral equivalente. Espécies pioneiras ou de ambientes
abertos de restingas constituíram 58% do total amostrado
neste estudo. Esta relativa pobreza em termos de espécies
pioneiras disponíveis na RBEPS pode estar relacionada ao
pequeno tamanho da clareira (estrada) que representa uma
porcentagem mínima em relação à área da floresta circunvizinha,
enquanto Sá (24) estima que 39% da mata foi perturbada e
esta mata também encontra-se isolada no meio de uma região
altamente degradada, com presença de grande número de espécies
pioneiras.
Deve-se levar em consideração
também que o cordão externo da RBEPS tem a sua idade avaliada
em 3.000 anos A.P. (1), enquanto o cordão interno, onde
está localizada a mata de Saquarema, tem sua idade atribuída
ao final do Pleistoceno, ou seja, mais de 10.000 A.P. (18).
A maior riqueza florística desta mata (114 espécies arbóreas
- Sá & Araujo, dados não publicados) pode estar relacionada
a este fato, além do menor grau de perturbação histórica
quando comparada com as matas da RBEPS, que foram bastante
modificadas pelos ciclos agro-econômicos que passaram pela
Ilha (2; 19).
Os resultados das análises
de solo tanto da área em regeneração quanto da mata fechada
encontram-se na Tabela 3. O valor de pH mais baixo encontrado
na mata pode ser explicado pela presença de 4,3 vezes mais
matéria orgânica no solo, o que pode provocar um aumento
de acidez. Uma densidade mais alta de raízes na mata também
pode contribuir para acidificação do solo por
meio da respiração e
ainda pela extrusão de prótons. Por outro lado, o poder
tampão dos solos varia diretamente com sua capacidade de
trocar cátions. Solos mais ricos em matéria orgânica são
mais resistentes à mudança de pH (14). Assim sendo, estes
solos teriam uma menor tendência à neutralização do que
os da estrada com relação à deposição de aerossóis marinhos.
TABELA 3: Valores
de fertilidade de solo (média de 6 repetições) até 20cm
de profundidade na área em regeneração e na mata de restinga
na Reserva Biológica Estadual da Praia do Sul. Valor S =
somatório das bases trocáveis (Ca, Mg, K e Na). Valor T
= capacidade de retenção de cátions pelo solo. T.F.S.A.
= Terra fina seca ao ar.
| |
Área
em regeneração |
Mata
intacta |
Coef.
Var. (%) |
| pH
em água |
4,90
* |
4,6
* |
- |
| Ca++
a |
0,47
* |
1,00
* |
55,0 |
| Mg++
a |
0,38
* |
0.83
* |
45,4 |
| Al
+++ a |
0,00
* |
0,05
* |
154,9 |
| H+
+ Al +++ a |
2,80
* |
6,37
* |
46,2 |
| P
b |
4,70
|
3,20
|
34,4 |
| K+
b |
18,2
* |
34,0
* |
26,4 |
| Na
+ b |
12,0
* |
34,0
* |
46,6 |
| C.
org. c |
0,67
* |
2,89
* |
52,8 |
| Mat.
Org. c |
1,15
* |
4,98
* |
52,8 |
| Valor
S a |
0,87
* |
2,15
* |
31,1 |
| Valor
T a |
3,36
* |
8,52
* |
36,0 |
* amostras estatisticamente diferentes
a nível de 1% pelo teste "t" de Student.
a meq/100 g T.F.S.A.
b ppm
c %
Os teores de Ca++, Mg++
e K+ apresentaram diferenças estatisticamente significativas
pelo teste "t" de Student (a = 0,01), ocorrendo, em todos
os casos, teores mais elevados na mata de restinga. Apesar
da mata conter um teor de 0,05 meq de Al+++/100g de T.F.S.A.,
este é considerado baixo demais para ocasionar distúrbios
fisiológicos nas plantas (14). A diferença de P no solo
não foi estatisticamente significativa nos dois locais,
mas deve ser destacado que grande parte do P do sistema
encontra-se estocado na vegetação de mata, cuja biomassa
é notavelmente mais alta. Quanto ao Na, o teor de 2,8 vezes
maior na mata pode ser atribuído à interceptação da salsugem
pelas folhas da vegetação da mata, que é muito mais densa
do que a da estrada. Clayton (9) afirma que os dosséis florestais
constituem-se em eficientes coletores de aerossóis, o que
contribui para aumentar o teor de Na no solo pela posterior
deposição de folhas e galhos. O teor de matéria orgânica
é 4,3 vezes maior na mata de restinga do que no solo da
área em regeneração. O valor baixo (1,15%) do solo desta
pode ser creditado à baixa produção de serapilheira pela
biomassa colonizadora do leito da estrada e as diferentes
taxas de decomposição. A serapilheira acumulada na área
em regeneração foi de 509±238g.m-2 e 920±170g.m-2 (peso
seco) na mata. Deve-se no entanto considerar que parte significativa
da serapilheira depositada no leito da estrada é proveniente
da mata contígua, cuja altura média (5,2m) e projeção de
suas copas sobre a estrada permite uma considerável taxa
de deposição de serapilheira alóctone sobre a mesma.
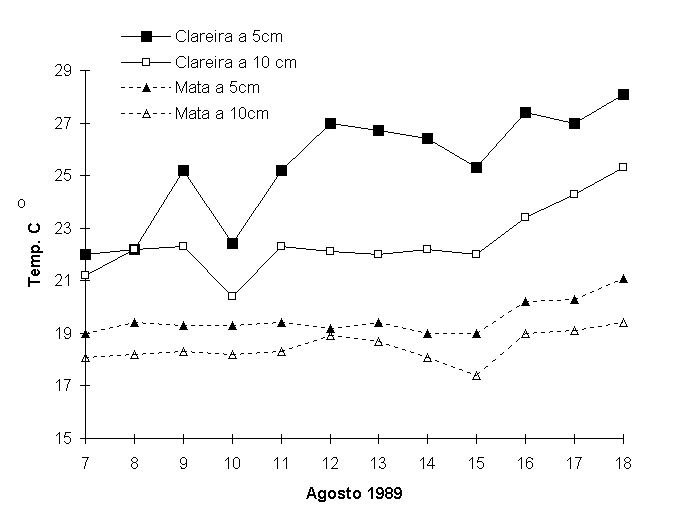
A Figura 4 apresenta
dados de temperatura do solo a 5 e 10cm de profundidade
na mata e na clareira, no período de 7 a 18 de agosto de
1989 às 14:00h. Deve-se destacar que o mês de agosto se
caracteriza por temperaturas mais baixas e portanto as diferenças
são menos significativas. A 10cm de profundidade, a variação
entre as temperaturas do solo da mata e da clareira são
menos intensas do que a 5cm. No entanto, é pouco representativa
a diferença da temperatura do solo da mata entre 5 e 10cm.
Isto pode ser atribuído ao papel do dossel da mata conservada
que reduz as variações diuturnas sobre a superfície do solo.
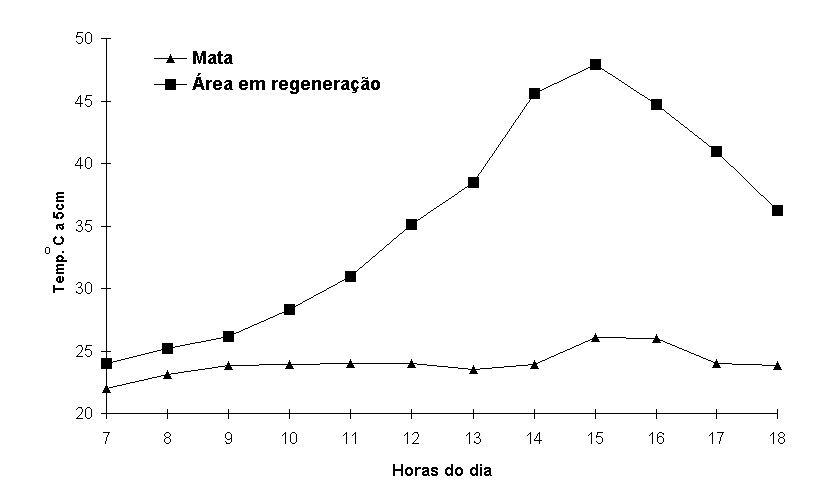
A Figura 5 mostra os
valores de temperatura do solo a 5cm tomados de hora em
hora no dia 8/01/91. Enquanto que a temperatura na mata
de restinga manteve-se em uma faixa de variação de 4,2oC,
a da clareira atingiu uma variação de 24oC. O pico de temperatura
alcançado no dia 8 de janeiro (48oC) deve ser ultrapassado
em fevereiro, em que a temperatura do ar atinge os valores
mais altos do ano. Esta variação de temperatura da clareira
representa um fator de inibição de numerosos processos biológicos
(27).
Figura 4.
Temperatura do solo a 5 e 10cm de profundidade na mata e
na área em regeneração entre 7 e 18 de agosto de 1989, ás
14h, na Reserva Biológica Estadual da Praia do Sul, Ilha
Grande, RJ.
Figura 5.
Marcha da temperatura do solo a 5cm no dia 8 de janeiro
de 191 na mata de restinga e na área em regeneração, na
Reserva Biológica Estadual da Praia do Sul, Ilha Grande,
RJ
Muito possivelmente
estes fatores, em conjunto com a temperatura do solo constituem
os principais determinantes da composição florística e estrutura
da vegetação da área em regeneração. As condições abióticas
ditadas pela natureza do distúrbio podem estar contribuindo
não só para a baixa biomassa de indivíduos colonizadores
do leito da estrada, como também para a baixa diversidade
florística, visto que apenas duas espécies (Byrsonima
sericea DC. e Cecropia lyratiloba Miq.)
representam 60% do VI. Ainda a respeito da baixa diversidade
florística, é de se considerar que os fatores abióticos
(principalmente temperatura e disponibilidade de água) podem
estar representando um impedimento à germinação de sementes
que chegam ao leito da estrada por dispersão, seja esta
por qualquer forma que ocorra (anemocoria ou zoocoria) ou
ainda, por queda direta dos frutos da mata sobre a estrada.
É de se esperar que o número de propágulos que a atinge
seja elevado. Por outro lado, a vegetação da estrada encontra-se
numa dinâmica sucessional caracterizada por uma baixa mortalidade
de indivíduos (27 indivíduos mortos/ha). A mata contígua
apresenta uma média de 213 indivíduos mortos/ha, o que sugere
que esteja em um estágio sucessional mais climáxico (Tabela
1).
Os resultados encontrados
mostram que as restingas constituem um ecossistema de regeneração
muito lenta se comparado à mata de encosta (10) levando-se
em consideração o número de espécies surgidas, seu arranjo
espacial e, ainda, o intervalo de tempo (8 anos). Intervenções
antrópicas neste ecossistema acarretam, portanto, distúrbios
não superáveis a médio prazo.
AGRADECIMENTOS
Agradecemos a Miriam
Cristina Alvarez-Pereira e Cyl Farney Catarino de Sá pela
ajuda indispensável nos trabalhos de campo.
ABSTRACT
Vegetation structure
and edaphic factors in a disturbed area of coastal plain
forest in the Praia do Sul Biological Reserve, Rio de Janeiro,
Brazil. A stretch of sandy coastal plain forest in Rio
de Janeiro State (Praia do Sul Biological Reserve) was disturbed
in 1980 when several roads were opened up and then abandoned.
Vegetation structure was analyzed 8 years after the disturbance.
All trees and shrubs (dbh ³ 2.5cm; height ³ 2m) were sampled
in 30 quadrats (5 x 5m) randomly located along the road
that runs parallel to the beach. Accumulated litter, soil
fertility and soil temperature were analyzed in the disturbed
area and in the adjacent forest. Only 18 species were found
in the disturbed area, the most important being Byrsonima
sericea DC. followed by Cecropia lyratiloba Miq.
and C. glazioui Snethl. Accumulated litter in the
disturbed area was 509 ± 238g.m-2 versus 920 ± 170g.m-2
in the forest. Organic matter and soil nutrient levels (excluding
phosphorous) were greater in the forest. Soil temperature
fluctuations were greater in the disturbed area. Forest
soils retained twice the amount of cations as disturbed-area
soils. Low organic matter content (1.15%) in the soils of
this area is a possible inhibiting factor of vegetation
growth. Although no further disturbance took place during
this 8-year period and the adjacent forest consitutes a
rich source of propagules, it appears that regeneration
of sandy coastal plain forest is a very slow process.
Key words: coastal
forest, disturbance, edaphic factors, biological reserve,
Brazil.
REFERÊNCIAS BIBLIOGRÁFICAS
- Amador, E.S. Geologia e geomorfologia
da planície costeira da Praia do Sul - Ilha Grande - uma
contribuição a elaboração do plano diretor da Reserva
Biológica Estadual da Praia do Sul. Anuário Inst. Geociências
UFRJ: 35-56, 1988.
- Araujo, D.S.D. & R.R.Oliveira.
Reserva Biológica Estadual da Praia do Sul (Ilha Grande,
Estado do Rio de Janeiro): lista preliminar da flora.
Acta bot. brasil., 1(Supl.):83-94, 1988.
- Araujo, D.S.D. & A.L.Peixoto.
Renovação da comunidade vegetal de restinga após uma queimada.
In: Anais Congresso Nacional Botânica, 26º, Acad.
Brasil. Ciências, Rio de Janeiro:1-17, 1977.
- Bazzaz, F.A. & S.T.A.Pickett.
Physiological ecology of tropical succession: a comparative
review. Ann. Rev. Ecol. Syst., 11:287-310, 1980.
- Brower, J.E. & Zar, J.H.. Field
and laboratory methods for general ecology. Wm. C.
Brown Publ., Dubuque, Iowa. 1984.
- Brokaw, N.V.L. & Scheiner, S.M..
Species composition in gaps and structure of a tropical
forest. Ecology, 70:538-541, 1989.
- Chapman, H.D. & P.F.Pratt. Métodos
de analises para suelos, plantas y aguas. Ed. Trillos,
Mexico, 1973.
- Clark, A.C. & D.B.Clark. Análise
de la regeneración de árboles del dosel en bosque muy
húmido tropical: aspectos teóricos y praticos. Rev.
Biol. Trop., 35 (supl. 1): 41-54, 1987.
- Clayton, J.L. Salt spray and mineral
cycling in two California coastal ecosystems. Ecology,
53:74-81, 1972.
- Delitti, W.B.C. Estudos de ciclagem
de nutrientes: instrumentos para a análise funcional de
ecossistemas terrestres. Oecologia brasiliensis,
1: 470-485, 1995.
- Denslow, J.S.. Tropical rain forest
gaps and tree species diversity. Ann. Rev. Ecol. Syst.,
18:431-451, 1987.
- EMBRAPA/SNLCS. Maual de Métodos
de Análise de Solo. Rio de Janeiro. 1979.
- Franco, A.C., D.M.Valeriano F.M.Santos,
J.D.Hay, R.P.B.Henriques & R.A.Medeiros. Os micro-climas
das zonas de vegetação de praia da restinga de Barra de
Maricá, Rio de Janeiro. In: Restingas: origem, estrutura,
processos. CEUFF, Niteroi. Lacerda, L.D. et al. (orgs.),
pp. 413-425, 1984.
- Moniz, A.C. Elementos de Pedologia.
Polígono, Ed. USP, São Paulo, 1972.
- Hartshorn, G.S.. Gap-phase dynamics
and tropical tree species richness. In: Tropical
Forests. Acad. Press, London. Holm-Nilsen, L.B.
et al. (eds.), pp. 65-73, 1989
- Hartshorn, G.S.. Application of gap
theory to tropical forest management: natural regeneration
on strip clear-cuts in the Peruvian Amazon. Ecology,
70:567-569, 1989
- Hay, J.D. & L.D.Lacerda. Ciclagem
de nutrientes no ecossistema de restinga. In: Restingas:
origem, estrutura, processos. CEUFF, Niteroi. Lacerda,
L.D. et al. (orgs.), pp. 461-477, 1984
- Martin, L. & Suguio, K.. Excursion
route along the Brazilian coast between Santos (State
of São Paulo) and Campos (Northern State of Rio de Janeiro).
International Symposioum on Global Changes in South America
during the Quaternary: past - present - future. São Paulo.
Special Publ. no. 2. 1989.
- Oliveira, R.R. & Coelho-Neto,
A.L. O rastro do homem na floresta: a construção da paisagem
da Reserva Biológica Estadual da Praia do Sul (Ilha Grande,
RJ) a partir das intervenções antrópicas. Albertoa,
4(10): 109-116, 1996.
- Orians, G.H.. The influence of tree-falls
in tropical forests in tree species richness. Trop.
Ecol., 23: 255-279, 1982.
- Poulson T.L. & Platt, W.J.. Gap
light regimes influence canopy tree diversity. Ecology,
70:553-555, 1989.
- Richards, P.W. The Tropical Rain
Forest. Cambridge University Press, New York. 1952.
- Rizzini, C.T. Tratado de fitogeografia
do Brasil. Ed. USP, São Paulo. 1979.
- Sá, C.F.C. Estrutura e composição
de uma área perturbada de mata de restinga na Reserva
Ecológica Estadual de Jacarepiá, Mun. Saquarema, RJ.
Tese de Mestrado, Pós-graduação em Botânica, Museu Nacional,
UFRJ, 167 p, 1993.
- Uhl, C., K. Clark, N.Dezzio &
P.Maquireno. Vegetation dynamics in Amazonian treefall
gaps. Ecology, 69:751-763, 1988.
- Whitmore, T.C. Canopy gaps and the
two major groups of forest trees. Ecology, 70:
536-538, 1989.
- Whitmore, T.C. Tropical rain forests.
Clarendon Press, Oxford, 1990.
|
|
|