Voltar
LOCAL TOPOGRAPHY AND UNDERSTORY BIRD DISTRIBUTION
IN A CENTRAL AMAZONIAN SITE, BRAZIL.
Sérgio Henrique Borges 1* André Carvalhaes
2 Luiza Magali Henriques 3
1 Fundação Vitória Amazônica,
Manaus, Amazonas; 2 Universidade Estadual Paulista - Campus
de Botucatu, Botucatu, São Paulo; 3 Museu Paraense
Emílio Goeldi, Belém - Pará
--------------------------------------------------------------------------------
ABSTRACT
To investigate the influence of local
topography on the understory bird community in the "terra
firme" forest of the Central Amazon (Jaú National
Park), we captured birds with mist nets at different topographic
sites: upland and near stream sites. In 1680 net hours
we captured 461 individuals of 64 species at seven study
sites. More species and individuals were captured in the
uplands, but the difference was not significant. Polar
ordination showed that the two topographic levels were
quite distinct in terms of bird species composition. Some
species seemed to show habitat preference related to local
topography, but the number of captures was too small for
statistical analyses. The only exception was the species
pair Hylophylax poecilinota and H. naevia. The antbird
H. poecilinota was captured more often in upland (n= 20
captures) than near stream sites (n= 3 captures) and the
congeneric H. naevia was more captured near stream sites
(n= 22 captures) than in upland (n= 6 captures). Even
with small samples and short duration this study suggests
that local topography can affect the understory bird distribution
in "terra firme" forest.
Key words: Amazon birds, topography, vegetation structure,
Hylophylax naevia, Hylophylax poecilinota.
--------------------------------------------------------------------------------
INTRODUCTION
Previous studies in tropical areas have
demonstrated the importance of local topography for habitat
use by birds, both for individual species (4) as well
as for the entire community (2). At the local level, topography
can affect the organisms distribution by influencing the
understory microclimate, resource availability and soil
type (2,10).
Here we present the results of a short-term
study on the influence of local topography in the understory
bird community in a Central Amazonian site. Specifically
we asked the following questions: Does the number of species
and individuals captured in mist nets differ between sites
located in upland and near stream habitats? Is the species
composition affected by local topography? Which species
show association with a particular topographic level?
MATERIAL AND METHODS
Study sites
We selected seven independent sites
to sample birds distributed in two regions of "terra
firme" forest separated by more than 100km in Jaú
National Park: three sites near Miratucu Lake (61o 49’
W and 1o 57’ S) and and four in the Tambor (62o
26’ W and 2o 14’ S), a small village at middle
Jaú river. Jaú National Park (JNP) includes
an area of 2,272,000ha and is one of the largest tropical
forest reserves in the world. The major habitats in JNP
include unflooded "terra firme" forest and forest
inundated by black water rivers (igapó forest).
The rainfall in the region is seasonal with the dry season
extending from June to December and the wet season from
February to June. The water level in rivers and streams
in JNP also varies seasonally with low waters occurring
from September to January and flooded periods from March
to August. The field work was conducted in March, April
and June of 1995.
Two habitat types were sampled at each
study site: plateau (PS) and near stream (NS). The plant
species composition differs between these topographic
levels (Leandro V. Ferreira, pers. comm.). Some palm species
(especially Jessenia pataua) and thickets as Heliconia
spp. were more common in near stream sites. In some places
the streams were dry during the sampling period.
Field procedures
We sampled birds with mist nets (12x2m
and 36mm mesh) with one net-line of 10 nets in each habitat
type. The two habitat types were sampled simultaneously.
The sampling effort was identical in these two topographic
levels. The PS and NS sites were separated by distances
varying between 30 and 50 meters. The net-lines were open
between 6:00 AM and 12:00. Each site was sampled on two
consecutive days; all birds captured were banded.
For the description of the vegetation
structure of study sites, four measurements of canopy
cover were taken on each side of every net using a spherical
densiometer.
Data analysis
Because the number of captures differed
between habitats we compared species richness between
PS and NS sites by a rarefaction method. Rarefaction generates
an expected number of species in a sample with standard
size using the following formula (3):
E(Sn)= å { 1- [ (N-Ni/ n) / (N/n)]
}
where:
E(Sn) = The number of species expected
in a standardized sample
n = The standard size of the sample
N = The total number of individuals in
the sample
Ni = The number of individuals of each
species i in the sample.
Bray Curtis polar ordination based on
Sorensen’s similarity coefficient as distance measure
was undertaken to compare species assemblages between
the study sites. As there are larger numbers of zeros
in the matrix species by sites we transformed the original
data using a Beals Smoothing that reduces the noise in
the data matrix (5). Species captured only once were not
considered in the ordination. Detailed descriptions of
this ordination method can be found in (3, 5). As a method
of capture, nets present problems including biological
and environmental variables that can affect the data interpretation
(1, 8). In order to minimize these problems, we performed
an ordination eliminating from the samples the species
highly mobile (hummingbirds, ant-followers and manakins)
which tend to have their abundance overestimated by mist
nets, and the species that tend to forage above the nets
(Appendix). This procedure has as a consequence a substantial
reduction of the sample size: 36% of species and 43% of
the captures were not considered.
RESULTS
In 1680 net hours we captured 461 birds
representing 64 species (Appendix). The cumulative curve
of species by captures showed that new species can be
added to local lists with increased sampling effort (Figure
1). However, field observations in the study sites both
by sight and song detection indicated that the nuclear
species (sensu 7) of the understory bird community were
sampled.
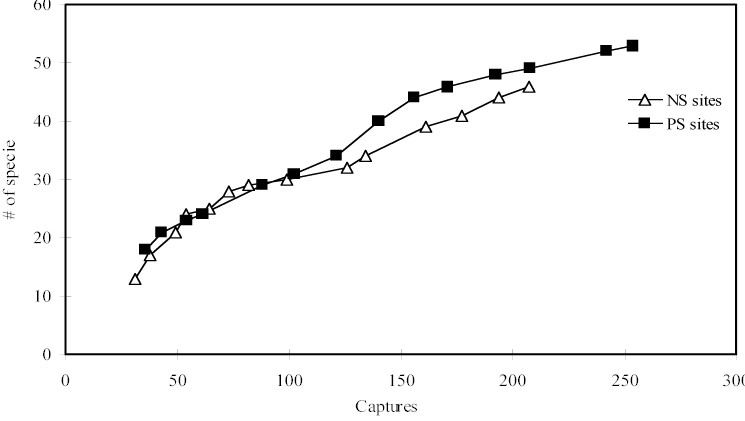
Figure 1 - Cumulative curve of species by captures in
near stream sites (NS) and plateau sites (PS).
Measurements of canopy cover as an index
of vegetation structure show that NS sites are more open
(mean = 15% of openness ± 6,52) than PS sites (mean
= 11,6 % ± 4,11) (Mann Whitney U-test, p < 0,05).
More species and individuals were captured
in PS than in NS sites (Table I), but the difference was
not significant (Mann Whitney U- test, p > 0,05). Similarly,
using a standard sample of 180 captures more species were
captured in PS sites, but the difference was very small
(Table I).
Table 1 - Sumary of the species richness and abundance
of birds in two topographic levels. PS are plateau sites
and NS are near stream sites.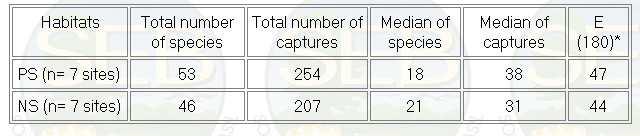
* Number of species expected by a standard sample of
180 captures.
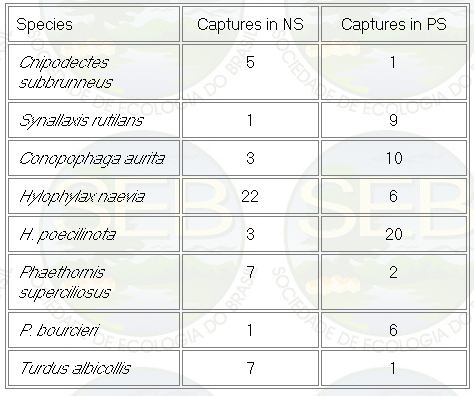
Appendix - Numbers of captures in NS and PS sites in
Jaú National Park. Asterisks indicate species not
considered in ordination analyses (see Methods).
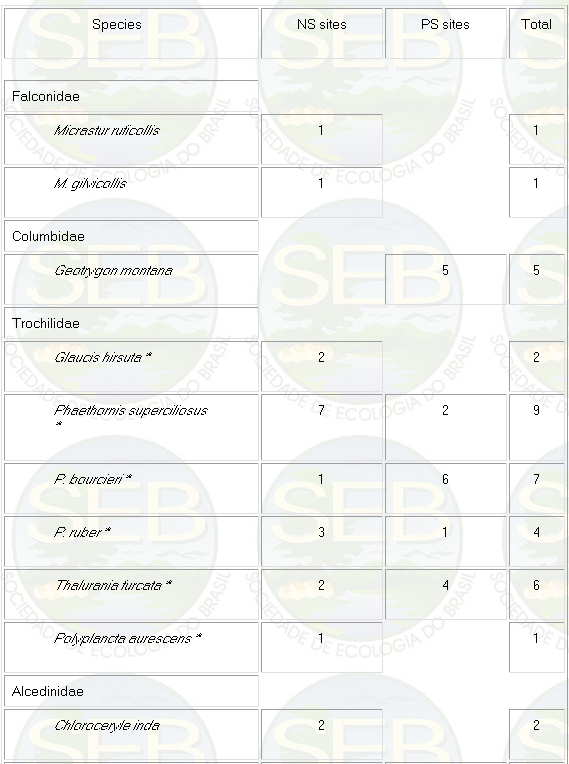
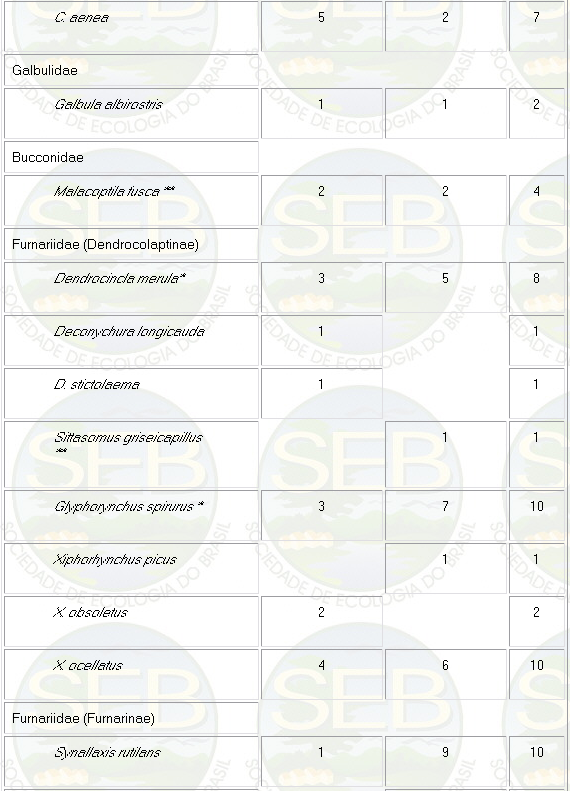
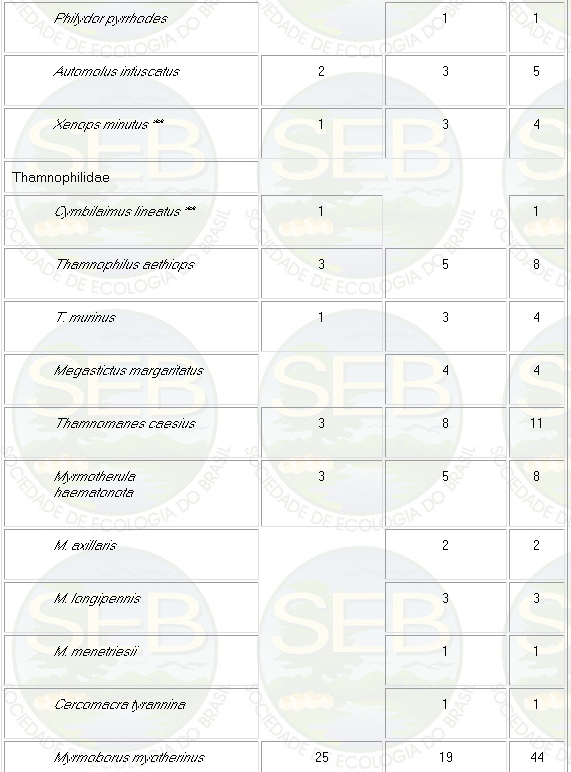
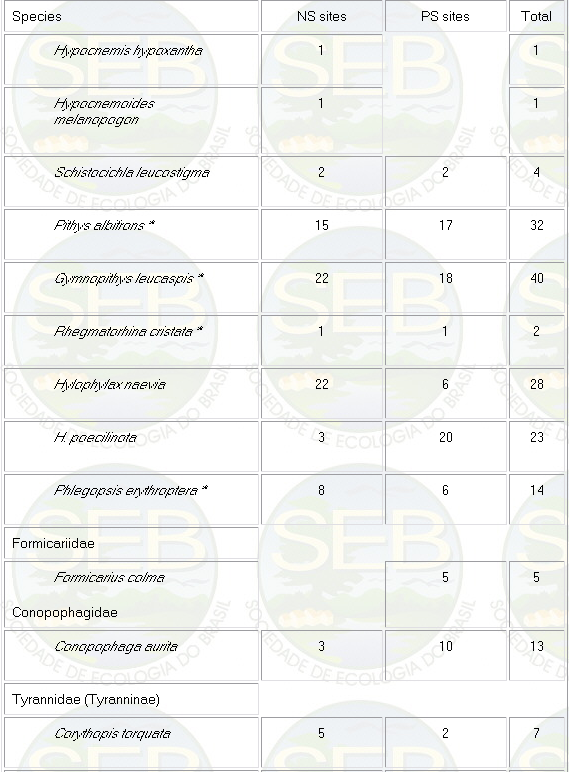
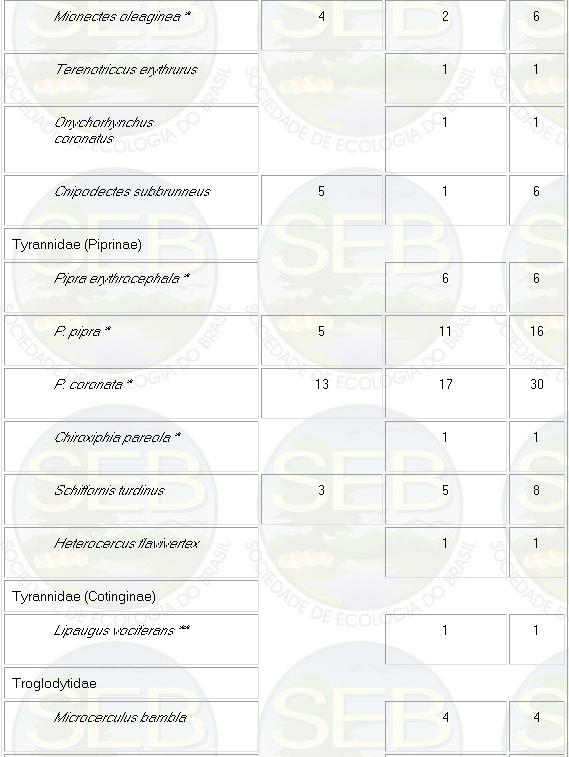
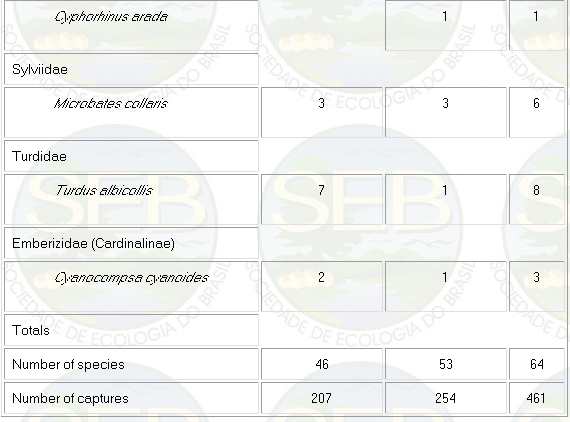
* Not included in analyses because spacing system highly
mobile (see Remsen 1996)
** Not included because foraging above net level.
Some bird species apparently showed
preference for habitats related to local topography (Table
II). Unfortunately, the number of captures was too small
to confirmed the statistical accuracy of these tendencies.
The only exception was the species pair Hylophylax poecilinota
and H. naevia with good sample size. H. naevia was significantly
more captured in NS than in PS sites (Mann Whitney U-test,
p < 0,05). On the other hand, H. poecilinota was more
abundant in PS sites (Mann Whitney U-test, p < 0,05).
Table 2 - Numbers of captures of some bird species with
some tendency in habitat preference. PS are plateau sites
and NS are near stream sites
The ordination results show that the
two topographic levels are quite distinct in species composition
(Figure 2a,b). When all species are analysed (Fig 2a)
the bird communities are clearly separated in two groups.
Similar results were obtained when some selected species
were eliminated from the samples (see methods) (Figure
2b). The ordinations suggested that net-lines placed in
PS and NS sites captured different "sub-groups"
of understory birds.
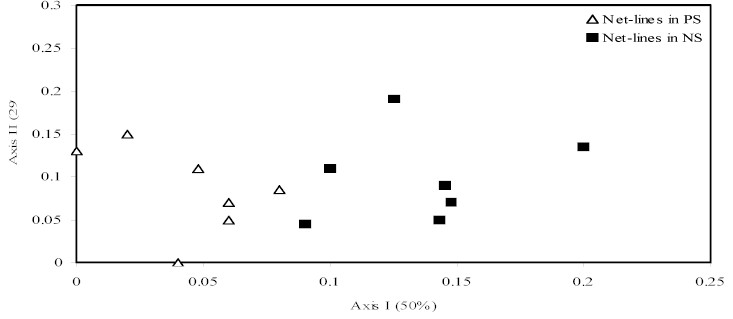
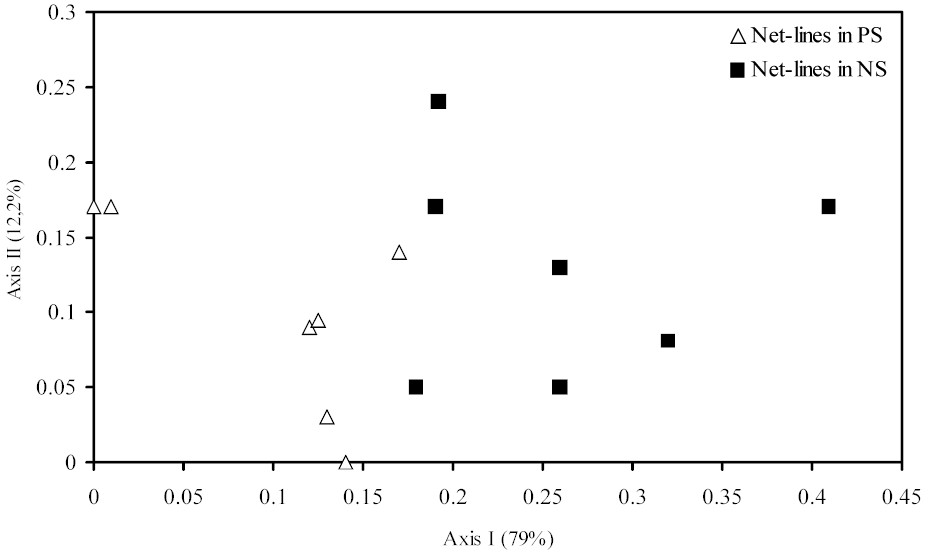
Figure 2 - Ordinations of the net-lines in near stream
(NS) and plateau sites (PS) including all species (a)
and excluding high mobiles species and those that foraging
above nets (b). In parenthesis are percents of variation
in the original matrix explained by each axis.
DISCUSSION
During this study no consistent effect
of local topography in the abundance and species richness
of understory birds was found. However, our results indicated
that two topographic levels contain distinct understory
bird communities. Habitat selection associated with local
topography also has been reported in plants (Melastomataceae
and Ferns - 10 and Palms - 6), frogs (9) and Basileuterus
warblers (4).
What environmental features related
to topography are important to understory birds? Classically
the vegetation structure is considered an important factor
in determining the habitat use by birds. Additionally,
it suggested that microclimate gradients of humidity,
clearly related to topography, can influenced habitat
selection directly through the impact on bird physiology
or indirectly in food resources availability (2). In fact,
activity patterns of birds are correlated with microclimate
gradients (2). We did not measure microclimate parameters
or food resources available for birds, but the vegetation
structure measured by canopy openness is different between
the two habitats sampled. This differences probably results
from the greater incidence of treefall gaps in NS sites.
The antbird H. naevia, one of the species with more tendency
to selected habitats in NS sites, was frequently observed
in gaps. On the other hand, H. poecilinota follow army
ants (11 and pers. obs.) and apparently avoid NS sites.
These results suggest that vegetation structure associated
with local topography can be an important factor in habitat
selection for understory birds.
ACKNOWLEDGMENTS
We thank Lindomar Alves Moreira for
help in field work with nets and Claude Gascon, Susan
Laurence and José Gomes for critical comments and
help in the English version. Field work was supported
by WWF (World Wildlife Fund), PNMA ( Plano Nacional do
Meio Ambiente), CAPES and CNPq (though the grant to the
senior author). This publication represents nº 3
of the Jaú Project technical series.
--------------------------------------------------------------------------------
RESUMO
Topografia local e a distribuição
de aves de sub-bosque num sítio da Amazônia
Central, Brazil. Para verificar a influência da
topografia local na comunidade de aves de sub-bosque numa
mata de terra firme da Amazônia Central (Parque
Nacional do Jaú) foram capturadas aves usando-se
redes em dois níveis topográficos: platôs
e baixios (locais próximos a igarapés).
Após 1.680 horas/rede foram capturados 461 indivíduos
de 64 espécies de aves em sete sítios de
estudo. Mais espécies e indivíduos foram
capturados nos platôs, mas a diferença não
foi significativa. Uma análise ao nível
de comunidade utilizando-se ordenação polar
parece demonstrar que os dois níveis topográficos
são distintos em termos de composição
de espécies. Algumas espécies parecem demonstrar
preferência de habitats relacionada à topografia
local, mas o número de capturas destas espécies
é muito pequeno para análises estatísticas.
A única exceção é o par de
espécies Hylophylax poecilinota e H. naevia. O
formicarídeo H. poecilinota foi mais capturado
em platôs (n= 20 capturas) do que em baixios (n
= 3 capturas), enquanto H. naevia foi mais capturado em
baixios (n= 22 capturas ) do que em platôs (n= 6
capturas). Mesmo com amostras pequenas e sendo de curta
duração, este estudo sugere que a topografia
local pode influenciar a distribuição de
aves de sub-bosque em matas de terra firme.
Palavras-chave: aves amazônicas, topografia, estrutura
da vegetação, Hylophylax naevia, Hylophylax
poecilinota
--------------------------------------------------------------------------------
REFERENCES
Karr, J. Surveying birds with mist nets.
Studies in Avian Biology 6: 62-67, 1981.
_____; Freemark, K. Habitat selection
and environmental gradients: dynamics in the "stable"
tropics. Ecology 64 (6): 1481-1494, 1983.
Ludwig, J.A. and Reynolds, J.F. Statistical
ecology - a primer on the methods and computing. Wiley
Interscience publication,1988, 337 pp.
Marini, M. A.; Cavalcanti, R.B. Habitat
and foraging substrate use of three Basileuterus warblers
from Central Brazil. Ornitologia Neotropical 4: 69-76,
1993.
McCune, B; Mefford, M.J. PC-ORD. Multivariate
analysis of ecological data. Version 2.0. MjM Software
Design, Gleneden Beach, Oregon, USA, 1995, 126 pp.
Peres, C. Composition, density, and fruiting
phenology of arborescent palms in an Amazonian terra firme
forest. Biotropica 26 (3): 285-294, 1994.
Remsen, J.V. Jr. Use and misuse of bird
lists in community ecology and conservation. Auk 111(1):
225-227, 1994.
_____________ Misuse of data from mist-net
captures to assess relative abundance in bird populations.
Auk 113 (2): 381-398, 1996.
Toft, C.A. Seasonal variation in populations
of Panamanian litter frogs and their prey: a comparison
of wetter and drier sites. Oecologia 47:34-38, 1980.
Tuomisto, H., Ruokolainen, K., Kalliola,
R. Linna, A., Danjoy, W.; Rodriques, Z.. Dissecting Amazonian
biodiversity. Science 269: 63-66, 1995.
Willis, E.O. The behavior of Scale-backed
Antbirds. Wilson Bull. 94(4): 447-462, 1982.
