Voltar
Impacto da variação
do nível de água no ciclo de vida da macrófita
aquática Pontederia cordata var. ovalis (Mart.)
Solms, em área alagável do Pantanal Mato-grossense.
Jerry Magno F. Penha1, * , Carolina J. da Silva1 &
Irineu Bianchini-Júnior2
1 Departamento de Botânica e Ecologia - Instituto
de Biociências - Universidade Federal de Mato Grosso
– 78060-900, Cuiabá - MT, Brasil. 2 Departamento
de Hidrobiologia - Universidade Federal de São
Carlos - Caixa Postal 676 - 13565-905, São Carlos
- SP, Brasil.
--------------------------------------------------------------------------------
RESUMO
Pontederia cordata é uma espécie
de macrófita aquática comum em áreas
alagáveis do Pantanal Mato-grossense. Três
populações da espécie foram estudadas
ao longo de 14 meses. O objetivo do estudo foi descrever
seu ciclo de vida em áreas alagáveis. Os
resultados indicam que características do ciclo
vital da espécie variam entre populações
que ocupam áreas submetidas a diferentes níveis
de estresse hídrico. Indivíduos que ocupam
áreas de fase seca acentuada são anuais.
Indivíduos que ocupam áreas de fase seca
moderada são perenes. Sugere-se que as populações
que habitam áreas alagáveis do Pantanal
dependem da variação do nível de
água para manterem sua diversidade genética
(regeneração por sementes).
Palavras chave: Pontederia cordata; ciclo de vida; Pantanal
Mato-grossense; áreas alagáveis; conceito
de pulso de inundação.
--------------------------------------------------------------------------------
INTRODUÇÃO
O Pantanal Mato-grossense é uma
extensa planície inundável. Em tais ambientes,
a enchente (pulso de inundação) é
encarada como a maior força seletiva agindo sobre
a biota (15). O caráter cíclico da inundação
engendra alternância sazonal entre a fase terrestre
e fase aquática em um mesmo sítio. A previsibilidade
dessa pressão ambiental induz o desenvolvimento
de mecanismos adaptativos nos organismos que ocupam tais
áreas. Adaptações relacionadas ao
pulso de inundação têm sido descritas
para vários organismos que habitam áreas
alagáveis (2, 9, 13, 14, 26).
Macrófitas aquáticas constituem
um dos principais componentes estruturais desses ecossistemas
(8). Segundo Crawley (7) organismos produtores podem enfrentar
períodos desfavoráveis, através de
estratégias que lhes possibilitem tolerar ou evitar
as pressões ambientais. Para as macrófitas
aquáticas, estratégias adaptativas para
tolerar o estresse, p. ex. tolerância a inundação
e a dissecação, plasticidade fenotípica;
e para evitá-lo, p. ex. resistência das sementes
e esporos a inundação e a dissecação,
têm sido descritas como formas de garantir a persistência
nesses ambientes (16, 18, 20).
Em ambientes sujeitos a inundação,
a melhor forma de um organismo maximizar sua aptidão
e garantir a persistência de seus genes é
adotar uma estratégia adaptativa do tipo r (16).
Todavia, dados empíricos que suportem esta hipótese
são escassos. Nesse sentido, este estudo visa avaliar
o ciclo de vida da macrófita aquática (P.
cordata), em áreas alagáveis do Pantanal
Mato-grossense.
MATERIAL E MÉTODOS
O Pantanal Mato-grossense localiza-se
entre os paralelos 16° e 22° S e os meridianos
55° e 58° W. Ocupa, em território brasileiro,
aproximadamente 44,21%, dos 380.000km2 da bacia do Alto
Paraguai (6). A região está sob influência
do sistema climático tropical seco e úmido
(24). O ciclo hidrológico é estacional (1,
6). A altitude média é inferior a 200m (3).
Solos hidromórficos predominam em 92% da área
(4).
O estudo foi realizado na Fazenda Ipiranga,
município de Poconé-MT (Figura 1). O local
situa-se entre os paralelos 16° 24’N e 56º40’W.
A região é formada por extensos campos inundáveis
entremeadas por vegetação de cerrado e mata
semicaducifólia. O estudo foi conduzido em campos
inundáveis.
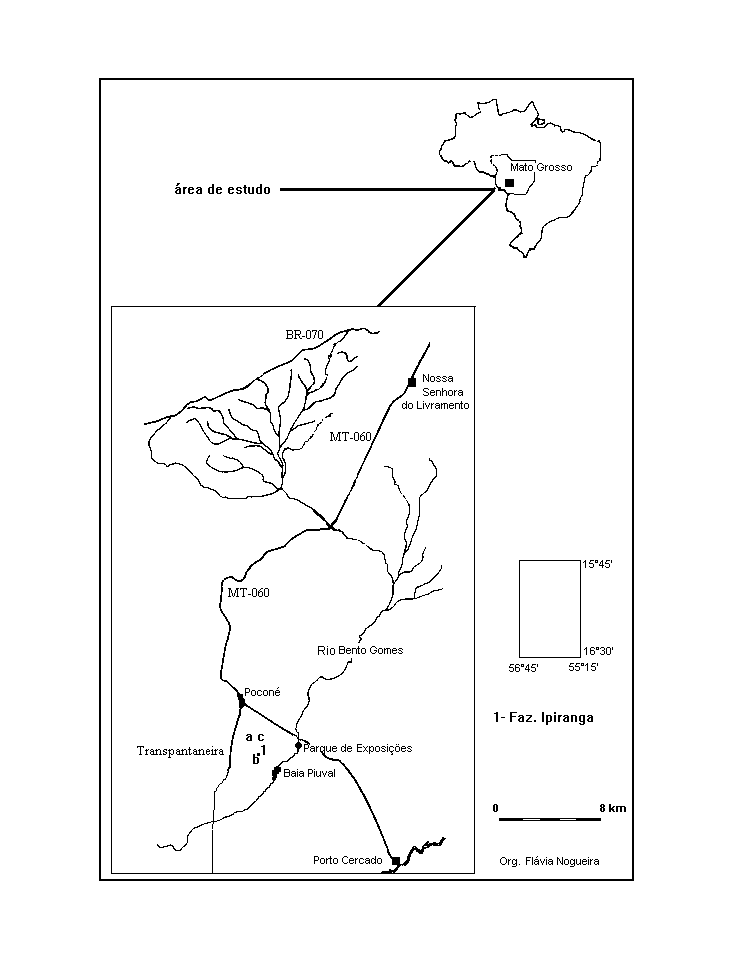
Figura 1 - Localização da área de
estudo, com indicação dos locais de amostragem
(A, B e C).
O trabalho foi desenvolvido em três
áreas (A, B, C) (Figura 1). Na área A, denominada
de área alagável de fase seca moderada,
a fase seca restringiu-se a dois meses (agosto e setembro).
A área permaneceu alagada durante onze meses, dos
quatorze de realização do estudo. O pico
da enchente ocorreu em março, 440mm medido na régua
(Figura 2). A área B apresenta características
hidrológicas semelhantes a área A, porém
com fase seca acentuada (seis meses), de meados de junho
a meados de dezembro. A área C localiza-se próximo
a área A. Todavia, sua maior profundidade, decorrente
de retirada de terra para construção de
estrada, possibilita permanência da lâmina
de água durante todo o ano, assemelhando-a a um
lago. Cabe ressaltar que o maior período de alagamento
da área A é decorrente do represamento por
uma estrada que corta o local. Desta forma, o padrão
hidrológico característico da região
é o da área B.
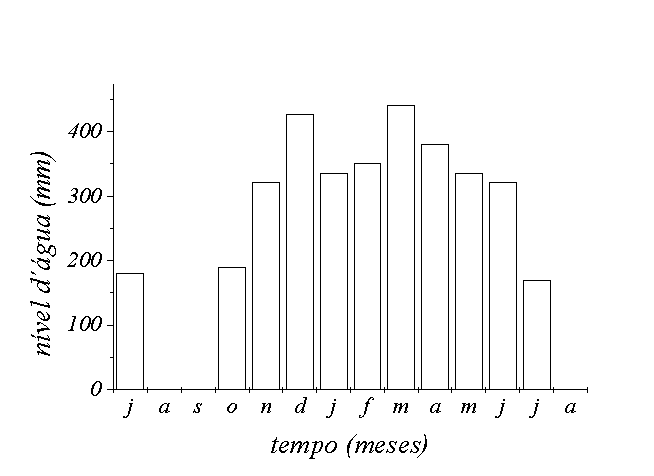
Figura 2 - Variação temporal da profundidade
da água para a área A. Fazenda Ipiranga,
Município de Poconé, MT.
Na área A foi delimitada uma parcela
de aproximadamente de 100x50m, na qual todas as coletas
foram realizadas. As observações do número
de indivíduos e de suas estruturas (folhas, rametes
e ramos reprodutivos) foram efetuados por meio de métodos
destrutivo e não destrutivo, entre julho de 1992
e agosto de 1993. O primeiro compreendeu a distribuição
periódica (entre agosto de 1992 e agosto de 1993)
de 10 ou 20 quadrados (cada um com 0,25m2) ao longo de
um transect de 50m e a coleta de todos os indivíduos
neles incluídos (22). O material colhido foi conduzido
ao laboratório, lavado em água corrente,
separado de acordo com as estruturas dos componentes e
determinado quantitativamente. Os rametes da espécie
por permanecerem unidos através de um rizoma curto,
possibilitou uma contagem simples. O método não
destrutivo compreendeu a observação de 27
indivíduos previamente marcados com fita plástica
colorida, distribuídos em 9 parcelas (3 plantas
por parcela), entre julho de 1992 e junho de 1993.
Nas áreas B e C foram realizadas
observações relacionadas a estratégia
de regeneração da espécie (sementes
ou rebrotamento). Para a área B o procedimento
consistiu em distribuir 10 quadrados ao acaso, cerca de
15 dias após o início da enchente, e realizou-se
a contagem de plântulas e rebrotamentos. Para a
área C, que possuíu cerca de 100m2, o procedimento
consistiu em monitorar, a cada 15 dias, o surgimento de
plântulas.
Os resultados apresentados foram obtidos
da área A, com algumas exceções que
serão destacadas.
RESULTADOS
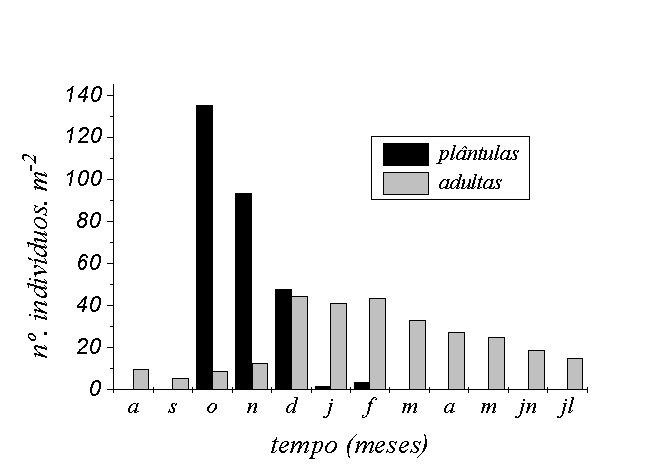
As populações estudadas
foram dependentes de seca periódica para recrutarem
suas plântulas. Na área A as plântulas
foram recrutadas nas primeiras semanas da enchente em
outubro (Figura 3A). Após esse pico da curva de
recrutamento, a densidade de plântulas tendeu a
diminuir. Novos recrutamentos ocorreram em menor escala
até o mês de janeiro (Figura 3 A). Na área
B, o recrutamento ocorreu com a chegada da enchente, no
mês de dezembro (densidade média de plântulas
no início da enchente = 114.m-2). Na área
C não houve recrutamento de plântulas (densidade
média de plântulas no início da enchente
= 0.m-2), apesar dos indivíduos terem produzido
sementes (observação pessoal do primeiro
autor).
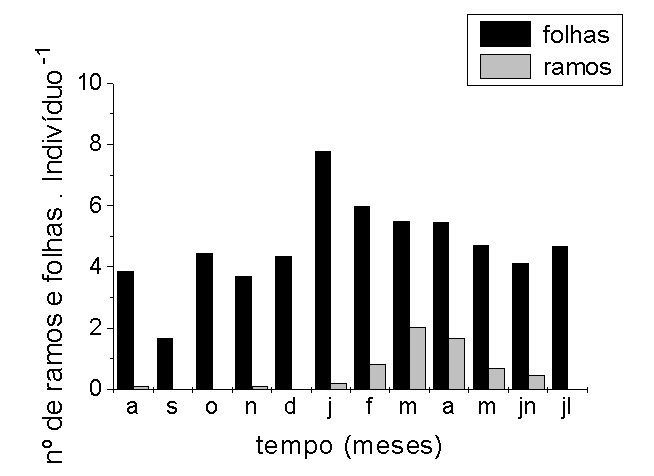
Os indivíduos alcançaram
maior densidade de folhas em janeiro (7,76 folhas por
indivíduo), cerca de 4 meses após o recrutamento
das plântulas (Figura 3 C).
A:
B:
C:
D:
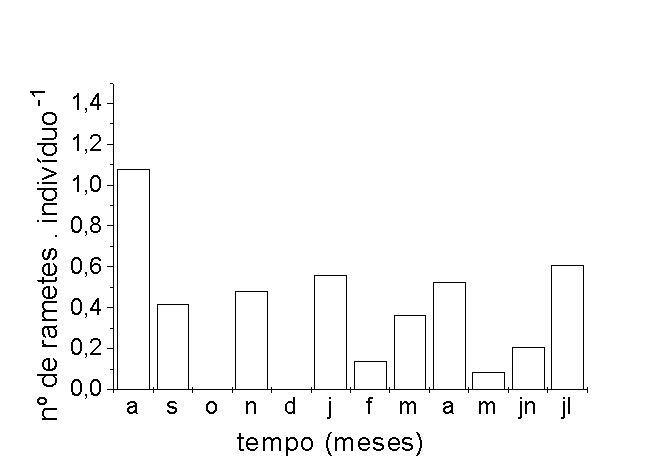
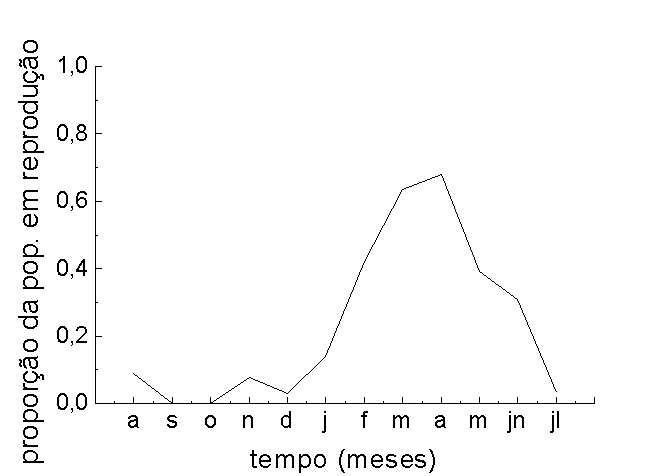
Figura 3 - Densidade média de P. cordata (plântulas
e adultas) na área de estudo (A), proporção
da população apresentando estruturas reprodutivas
(B), variação do número médio
de folhas e ramos reprodutivos por indivíduo (C)
e de rametes (D), no sítio amostral; Fazenda Ipiranga,
Município de Poconé, MT.
A população reproduziu
-se durante dez meses por ano. Indivíduos rebrotados
que sobreviveram à fase seca como os rizomas, começaram
a se reproduzirem em novembro, dois meses após
o início da enchente, originando um primeiro pico
de produção de ramos reprodutivos (Figuras
3 B e C e observação pessoal do primeiro
autor). Para indivíduos originados de sementes,
a floração iniciou-se na cheia, 4 meses
após a germinação. O pico de frutificação
ocorreu entre março e abril (Figuras 3 B e C).
Após esse pico de atividade reprodutiva, a fração
da população em reprodução
tendeu
a diminuir e, em junho, julho e agosto,
o número de ramos reprodutivos que não produziram
inflorescências (abortos) tendeu a aumentar (observação
pessoal do primeiro autor). As plantas produziram estruturas
reprodutivas, inclusive durante a fase seca, agosto (Figura
3 C). O aumento da densidade de ramos reprodutivos coincidiu
com diminuição da densidade de folhas (Figura
3 C).
O maior número de rametes por
indivíduo foi determinado na seca , setembro (Figura
3 D). É provável que o estresse hídrico
tenha sido o principal fator determinante da duração
de vida dos indivíduos das populações.
Na área A, os indivíduos apresentaram menor
número de folhas durante a fase seca, 1,65 folhas
por planta (Figura 3 C). Nesta, grande parte da população
secou e morreu. Alguns sobreviveram na forma de rizomas,
rebrotando com a chegada da enchente (densidade de indivíduos
rebrotados no início da enchente = 5.m-2 ). Todos
os indivíduos da área B morreram no decorrer
da fase seca. Esta conclusão baseou-se na ausência
de rebrotamentos no início da enchente da área
(densidade de indivíduos rebrotados no início
da enchente = 0.m-2 ). Não houve alteração
visível entre os adultos da área alagada
(observação pessoal do primeiro autor).
Assim, as populações das áreas A
e C foram compostas por indivíduos com ciclo de
vida perene. A população da área
B foi composta por indivíduos de ciclo de vida
anual.
DISCUSSÃO
P. cordata é uma espécie
de macrófita aquática distribuída
por todo o continente americano, de hábito perene
e comum em locais úmidos, próximos a rios,
riachos, lagos e charcos perenes (12). No Pantanal Mato-grossense
a espécie se reproduz por sementes e se propaga
vegetativamente. Neste caso os rametes formados (senso
25 e 11) permaneceram unidos pelo rizoma. O genete cresceu
em forma de roseta e teve uma estratégia exploratória
do tipo falange (phalanx) Esta estratégia foi associada
com a manutenção de bons sítios para
estabelecimento dos indivíduos (10).
A espécie foi tolerante à
variações de umidade, ocupando gradiente
de inundação, compreendendo ambientes permanentemente
alagados e periodicamente secos.
O ciclo de vida da população
foi ajustada a oscilação do nível
de água, e dependente de uma fase seca para se
renovar. Semelhante isto observou-se para Echynocloa polystachia,
um capim semi aquático que habita várzeas
da Amazônia Central (20, 21). Porém, E. polystachia
somente se reproduz por brotamento.
Há dois relevantes contrastes
entre investimento em reprodução por sementes
e propagação por botões germinativos,
que irão produzir os rametes (17). Recrutamento
a partir do banco de sementes permitiu a manutenção
da variabilidade genética na população,
enquanto que a partir de botões germinativos perpetuou-se
os genótipos experimentados e bem sucedidos. Com
base nestas observações supõe-se
que a modificação do regime hídrico,
através do impedimento da variação
do nível de água, determinou a redução
da diversidade genética da população.
Neste caso, é provável que haja extinção
da espécie, dependendo do tempo de viabilidade
das sementes armazenadas no banco de sementes. Segundo
Junk & Piedade (16), muitas espécies de herbáceas
que habitam planícies alagáveis do rio Amazonas,
próximo a Manaus, também são dependentes
da alternância entre a fase aquática e a
fase terrestre e "podem não ocorrer, se o
pulso de inundação for eliminado".
O padrão de reprodução,
com pico de produção de sementes na cheia,
pode maximizar a dispersão de sementes pela água
(20).
No Pantanal Mato-grossense observou-se
que P. cordata é uma espécie de hábito
de vida variável. Em áreas com fase seca
acentuada (área B), provavelmente o fator de mortalidade
seja independente do tamanho da planta, já que
devido a seca todos os adultos da área morreram.
Altas taxas de mortalidade de adultos favoreceu a seleção
de reprodução precoce (19). Esta determinou
um rápido desenvolvimento (5, 7, 19). As gerações
são discretas, e o hábito anual, semelparidade
e estratégia do tipo r foram favorecidos (Tabela
1).
Tabela 1 - Características do ciclo de vida da
macrófita aquática Pontederia cordata, em
relação a intensidade da estação
seca. Pantanal Mato-grossense, MT.
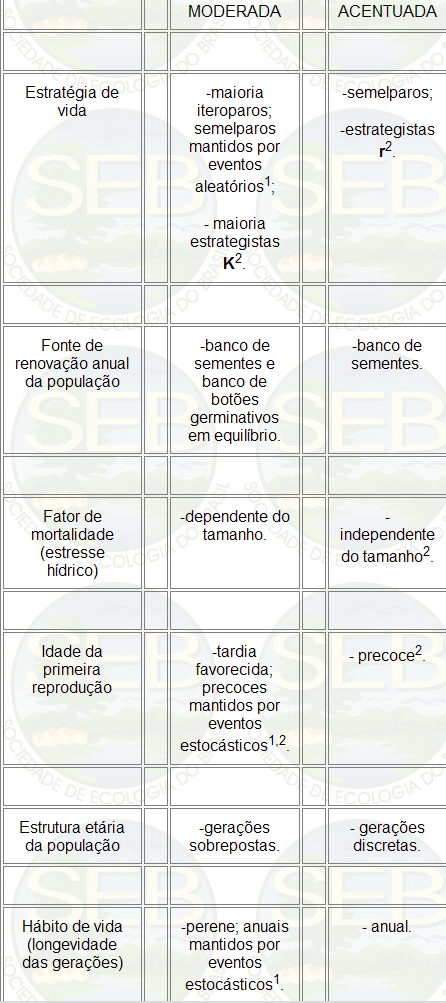
1 duração da fase seca.
2 traços hipotéticos baseados na literatura
(19, 23).
Indivíduos ocupando áreas
de seca moderada (área A) provavelmente foram sujeitos
a um fator de mortalidade dependente do tamanho. Tal afirmativa
é corroborada pelo fato da população
possuir tendência à translocar matéria
para estruturas subterrâneas no decorrer das fases
de vazante e seca (18). Indivíduos maiores poderiam
translocar mais materiais para os órgãos
de armazenamento e, provavelmente aumentar suas chances
de sobrevivência. Neste caso, a seleção
agiria favorecendo maior investimento na manutenção
e menor na reprodução (5, 7, 19) (Figura
3 C). O menor investimento em reprodução
no primeiro ano seria compensado pela garantia do sítio
de estabelecimento, maior tamanho e maior esforço
reprodutivo no segundo ano (5, 7). Nessas áreas,
a proporção de indivíduos recrutados
através de rebrotamento e a taxa de mortalidade
de plântulas devem ser variáveis, havendo
como conseqüência a sobreposição
de gerações (19). A seleção
agiria favorecendo, hábito perene, estrategistas
K e iteroparidade (Tabela 1).
Os resultados aqui apresentados sugerem
que a predição de que a melhor forma de
sobrevivência em áreas alagáveis seja
a estratégia adaptativa do tipo r, não é
aplicável para a totalidade do gradiente de inundação
que ocorre no Pantanal Mato-grossense.
AGRADECIMENTOS
Aos MS. Marcos E. Coutinho (EMBRAPA-CPAP)
e Soraia de A. Ferreira (Dep. Ecologia-UFMT) e aos revisores
anônimos, pela leitura e críticas ao manuscrito.
Ao Dr. Michael Schessl da Universidade de Ulm, Alemanha,
pelo auxílio na revisão da nomenclatura
da espécie estudada. À CAPES pela bolsa
de mestrado do primeiro autor. Ao senhor João Lousano
pelas facilidades fornecidas na área de estudo.
Contribuição número 20 do Projeto
Ecologia do Gran Pantanal (UFMT/FEMA/MPI), Programa SHIFT
(CNPq/ IBAMA/DLR); Cooperação Técnico-Científica
Brasil - Alemanha.
--------------------------------------------------------------------------------
ABSTRACT
The impact of water level variation on
life cycle of aquatic macrophytes Pontederia cordata var.
ovalis (Mart.) Solms, on floodplains of the Pantanal Mato-grossense.
Pontederia cordata is a common specie of aquatic macrophytes
in floodplains of the Pantanal Mato-grossense. Three populations
of the species were studied for 14 months. The aim of
this research was to describe its life cycle in floodplains.
The results indicate that characteristics of the life
cycle of the species varied between the populations that
occupied areas submitted to different level of hydric
stress. Individuals
that occupied areas in acute draught
phases are annuals. Individuals that occupied areas in
moderate draught phases are perennials. It is suggested
that the populations that inhabit floodplains in the Pantanal
depend on the variation of the water level to maintain
their genetic diversity (regeneration by seeds).
Key words: Pontederia cordata; life cycle; Pantanal Mato-grossense;
floodplains; flood pulse concept.
--------------------------------------------------------------------------------
REFERÊNCIAS
Adamoli, J.A. A dinâmica das inundações
no Pantanal in: Anais do primeiro simpósio sobre
recursos naturais e sócio-econômicos do Pantanal.
Embrapa-DDT, Brasília, 1986, p.63-76.
Adis, J.; Mahnert, V. On the natural history and ecology
of Pseudoscorpiones (Arachnida) from an Amazonian blackwater
inundation forest. Amazoniana, 9: 297-314, 1985.
Alvarenga, S.M.; Brasil, A.E.; Pinheiro, R.; Kux, H.J.H.
Estudo geomorfológico aplicado a bacia do Alto
Paraguai e
Pantanais Mato-grossenses. Boletim técnico
do Projeto Radambrasil (Série Geomorfologia, (1),
p.1-187, 1984.
Amaral Filho, Z. P. Solos do Pantanal Mato-grossense in:
Anais do primeiro simpósio sobre recursos naturais
e sócio-econômicos do Pantanal. Embrapa-DDT,
Brasília, 1986, p.91-103.
Begon, M.; Mortimer, M. Population ecology. A unified
study of animals and plants. Blackwell Scientific Publications,
Oxford, 1986, 220p.
Carvalho, N. O. Hidrologia da bacia do Alto Paraguai in:
Anais do primeiro simpósio sobre recursos naturais
e sócio-econômicos do Pantanal. Embrapa-DDT,
Brasília, 1986, p.43-49.
Crawley, M. J. Life history and environment in: Crawley,
M. J. (ed). Plant ecology. Blackwell Scientific Publications,
Oxford, 1986, p.253-290.
Esteves, F. A. Fundamentos de Limnologia. Interciência
- FINEP, Rio de Janeiro, 1988, 575p.
Gottsberger, G. Seed dispersal by fish in the inundated
regions of Humaita, Amazônia. Biotropica, 10: 170-183,
1978.
Harper, J.L. Population biology of plants. Academic press,
London, 1977, 892p.
Harper, J.L. The concept of population in modular organisms
in: May, R.M. (ed). Theoretical ecology: principles and
applications. 2ª ed., Blackwell Scientific Publications,
Oxford, 1981, p.53- 77.
Horn, C. N. Pontederiaceae. Conservatoire et Jardin botaniques
de la Ville de Genève & Missouri Botanical
Garden (Flora del Paraguay), Genebra, 1987, 28 p.
Irmler, U. Considerations on structure and function of
the central Amazonian inundation forest ecossistems with
particular emphasis on selected soils animals. Oecologia,
43: 1-18, 1979.
Irmler, U. Temperature dependant generationcycle for the
cicindelid beetle Pentacomia egregia Chaud. (Coleoptera,
Carabidae, Cicindelinae) of the Amazon valley. Amazoniana,
9: 431-439, 1985.
Junk, W. J.; Bayley, P.B.; Sparks, R.E.
The flood pulse concept in river-floodplain systems. Can.
Spec. Publ. Fish. Aquatic. Sci., 106: 110-127, 1989.
Junk, W. J.; Piedade, M.T.F. Herbaceous plants of the
Amazon floodplain near Manaus: species diversity and adaptations
to the flood pulse. Amazoniana, 12: 467-484, 1993.
Noble, J. C.; Bell, A. D.; Harper, J.L. The population
biology of plants with clonal growth - I. The morphology
and structural demography of Carex arenaria. Journal of
Ecology, 67: 983-1008, 1979.
Penha, J. M. F. Ecologia populacional de Pontederia lanceolata
Nuttal, em uma área alagável do Pantanal
Matogrossense - MT. São Carlos, 1994, 94 p (M.Sc.
Dissertation, Centro de Ciências Biológicas
e da saúde, UFSCar).
Pianka, E.R. On "r" and "K" selection.
Am. Nat., 104: 592-597, 1970.
Piedade, M. T. F. Biologia e ecologia de Echinochloa polystachya
(H. B. K.) Hitchcock (Graminae = Poaceae), capim semi-aquático
da várzea amazônica. Acta Limnol. Brasil.,
6: 173-185, 1993.
Piedade, M. T. F.; Junk, W. J.; Long, S.P. The productivity
of the C4 grass Echinochloa polystachya on the amazon
floodplain. Ecology, 72: 1456-1463, 1991.
Roberts, M. J.; Long, S.P.; Tieszen, L.L.; Beadle, C.L.
Measurementof plant biomass and net primary production
in: Coombs, J.; Hall, D.O.; Long, S.P.; Scurlock, J.M.O.
(eds). Techniques in bioproductivity and photosynthesis.
2ª ed., Pergamon press, Oxford, 1987, p.1-19.
Stearns, S.C. The evolution of life history traits: a
critique of theory and a review of data. Ann. Rev. Ecol.
Syst., 8: 145-171, 1977.
Tarifa, J. R. O sistema climático do Pantanal:
da compreensão do sistema à definição
de prioridades de pesquisa climatológica in: Anais
do primeiro simpósio sobre recursos naturais e
socio-econômicos do Pantanal. Embrapa-DDT, Brasília,
1986, p. 9-27.
White, J. The plant as a metapopulation. Ann. Rev. Ecol.
Syst., 10: 109-145, 1979.
Worbes, M. Structural and other adaptations to longterm
flooding by trees in central Amazonia. Amazoniana, 9:
459-484, 1985.
