
Voltar
A INFLUÊNCIA DA MACRÓFITA AQUÁTICA Echinochloa
polystachya (H.B.K.) HITCHCOCK NAS CARACTERISTICAS FÍSICAS
E QUÍMICAS DA ÁGUA NA ZONA DE DESEMBOCADURA DO RIO PARANAPANEMA
NA REPRESA DE JURUMIRIM - SP
Pompêo, M.L.M.1;
Henry, R.1; Moschini-Carlos1, V.;
Padovani, C.R.2
Depto. de Zoologia e 2
Bioestatística - UNESP - IB, - Botucatu - SP - 18618-000
RESUMO
Na zona de desembocadura
do rio Paranapanema na represa de Jurumirim (SP, Brasil)
a macrófita aquática emergente Echinochloa polystachya
(H.B.K.) Hitch. domina a fisionomia vegetal. Enraizada nas
margens, esta planta projeta seus colmos em direção à região
pelágica. Os padrões sazonais de variáveis físicas e químicas
da água no estande dessa gramínea e na calha principal do
rio foram estudados. O trabalho visa mostrar a influência
do estande de E. polystachya sobre essas variáveis.
O padrão de variação anual da temperatura da água e da concentração
de oxigênio dissolvido observada é decorrente das alterações
da temperatura do ar. As modificações na concentração de
material em suspensão e na condutividade elétrica foram
associadas à alternância do regime de precipitação durante
o ano. O rio apresentou tendência à homogeneidade nas variáveis
físicas e químicas devido à movimentação da massa dágua.
No estande, a camada de água mais próxima da superfície
apresentou características diferentes das zonas intermediária
e profunda. Comparando-se os valores de temperatura e de
concentração de oxigênio dissolvido na água do estande com
aqueles determinados no rio, as diferenças encontradas são
estatisticamente significativas. Foram atribuídas à presença
do estande de E. polystachya, que atua como barreira
física à passagem da água e à ação do vento, que reduz a
renovação e a homogeneização da massa de água.
Palavras-chave:
macrófita aquática, Echinochloa polystachya, represa,
rio Paranapanema, ciclo hidrológico, características físicas
e químicas
INTRODUÇÃO
Em regiões tropicais,
as macrófitas aquáticas presentes na zona de desembocadura
dos rios nos reservatórios apresentam grande biomassa, contribuindo
sobremaneira para a produção de matéria orgânica para o
ecossistema aquático. Podem apresentar importante papel
na transferência de nutrientes entre a fase aquática e a
terrestre (14), com a potencialidade de controlar a dinâmica
de nutrientes no ecossistema (21). Além disso, os estandes
das macrófitas aquáticas também possibilitam a existência
de inúmeros microhabitats, onde uma considerável gama de
organismos pode passar parte ou todo seu ciclo de vida,
depositando ovos, protegendo-se de predadores, alimentando-se.
Assim, a água do estande de macrófitas aquáticas poderá
ser um compartimento diferenciado das águas abertas, refletido
por diferenças físicas, químicas e biológicas. Os experimentos
efetuados por Dale & Gillespie (5), sugerem que a temperatura
da água sofre influências do próprio arranjo das folhas
da população de macrófitas aquáticas bem como do índice
de área foliar. A interceptação da energia luminosa pela
superfície foliar aquece a água localmente, enquanto que
na parte de baixo, devido ao sombreamento, tende a ser menor.
Desta forma, segundo Esteves (7), os estandes das macrófitas
aquáticas constituem-se num compartimento muito complexo,
que devem ser analisados separadamente da região de águas
abertas.
Este trabalho mostra
os padrões sazonais de variáveis físicas e químicas da água
no estande de Echinochloa polystachya (H.B.K.) Hitchcock
e na calha do rio Paranapanema (SP, Brasil). Visa também
verificar a influência do estande sobre essas variáveis.
LOCAL DE ESTUDO
A represa de Jurumirim
(reservatório Engenheiro Armando Avellanal Laydner) está
localizada na porção sul do Estado de São Paulo e é a primeira
represa no sentido montante-jusante do complexo de barragens
que se sucedem no rio Paranapanema. Na cota 570m, a água
de superfície do reservatório cobre uma área de cerca de
484km2 e o seu volume é estimado em 7.941hm3.
A bacia hidrográfica apresenta cerca de 17.800km2.
É um reservatório oligotrófico, com baixa produtividade
primária fitoplanctônica na zona de barragem, e o crescimento
das algas provavelmente é limitado pelo fósforo (9, 10).
Na zona de desembocadura
do rio Paranapanema no reservatório (Figura 1a), são observadas,
além do canal do rio, extensas áreas inundadas, ambas margeadas
por estandes de macrófitas aquáticas emergentes, predominantemente
por E. polystachya. A gramínea
E. polystachya apresenta um colmo robusto com mais
de 15m de comprimento, que está enraizado nas margens, projetando-se
em direção às águas abertas (Figura 1b). Esta macrófita
aquática apresenta elevada biomassa (1.150 a 2.756gPS/m2)
e estoque de nutrientes (0,36 a 4,43kgP/m2 e
9,6 a 46,0kgN/m2), e tem uma produção primária
estimada em 30t/ha/ano (21).
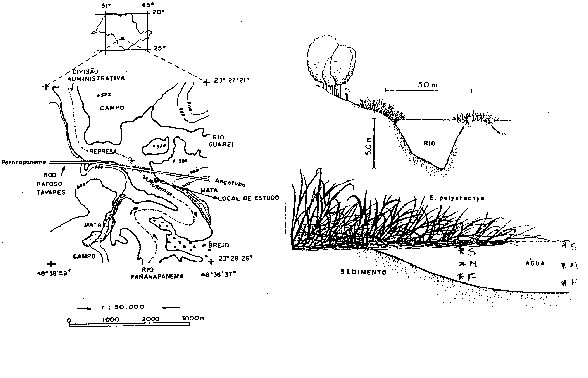
Figura 1: a)
Local de estudo na zona de desembocadura do rio Paranapanema
na represa de Jurumirim e b) estande de E. polystachya
com a localização das profundidades de colata de água. (S=
superfície; M= meio; F= fundo).
MATERIAIS E MÉTODOS
As amostras de água
foram coletadas mensalmente em 3 profundidades em dois pontos
fixos, um no interior do estande de E. polystachya e
outro imediatamente na margem do estande, no canal principal
do rio Paranapanema, (Figura 1).
Foram determinadas as
temperaturas do ar e da água com um termistor Toho Dentan
(± 0,10C), o material em suspensão total (MST)
(24, 29), o oxigênio dissolvido pelo método Winkler (8),
a condutividade elétrica (com um condutivímetro Hach Mod.
2511, tendo os valores corrigidos para 250C (8),
e o pH (potenciômetro Micronal B 371). Para a estimativa
dos teores dos nutrientes dissolvidos, a água foi previamente
filtrada em filtro Millipore AP40 e o filtrado armazenado
em frascos de polietileno e estocados em freezer. Em laboratório,
foram feitas as análises de nitrato e nitrito (17), amônio
(15), fosfato inorgânico dissolvido (FID) e fosfato total
dissolvido (FTD) (23).
A variação do nível
da água foi determinada mediante leitura em régua graduada
instalada no interior do estande. Foram obtidos os valores
das cotas diárias do nível da água (cotas das 9h00min) e
das vazões no setor operacional da barragem, as temperaturas
máxima e mínima e as precipitações diárias na estação climatológica
da Cooperativa Agro Industrial Holambra (Fazenda Holambra,
Paranapanema, SP), localizada a cerca de 30Km em linha reta
do local de estudo.
Os dados climatológicos
foram submetidos a uma análise de agrupamento através da
classificação hierárquica ascendente, utilizando-se a distância
Euclidiana, tendo como critério de agregação as médias das
distâncias ponderadas. Os cálculos foram efetuados através
do programa estatístico STAT-ITCF 4.0. Os dados foram previamente
padronizados pela subtração da média e divisão pelo desvio
padrão (4).
As diferenças entre
as profundidades de coleta de água (no rio e estande) em
cada um dos grupos experimentais (variáveis) foram detectadas
através do uso da técnica de análise multivariada, complementada
com a construção dos intervalos de confiança simultâneos
(18).
Os valores das variáveis
físicas e químicas na água do rio e do estande foram comparados
entre si e também com aqueles encontrados por outro autor
(19) em uma baía permanentemente inundada e ligada ao rio
Paranapanema, próximo ao local deste estudo, igualmente
através do método de Morrison (18).
RESULTADOS
As maiores temperaturas
do ar foram observadas no período de outubro/93 a março/94
(Figura 2). A variação anual da temperatura do ar medida
no local de estudo nos dias de coleta mostra padrão semelhante
à temperatura do ar medida no posto de observação climatológica
(Figura 3). A precipitação mostrou marcado padrão sazonal,
com maiores valores em janeiro de 94 e em setembro/93 (Figura
2).
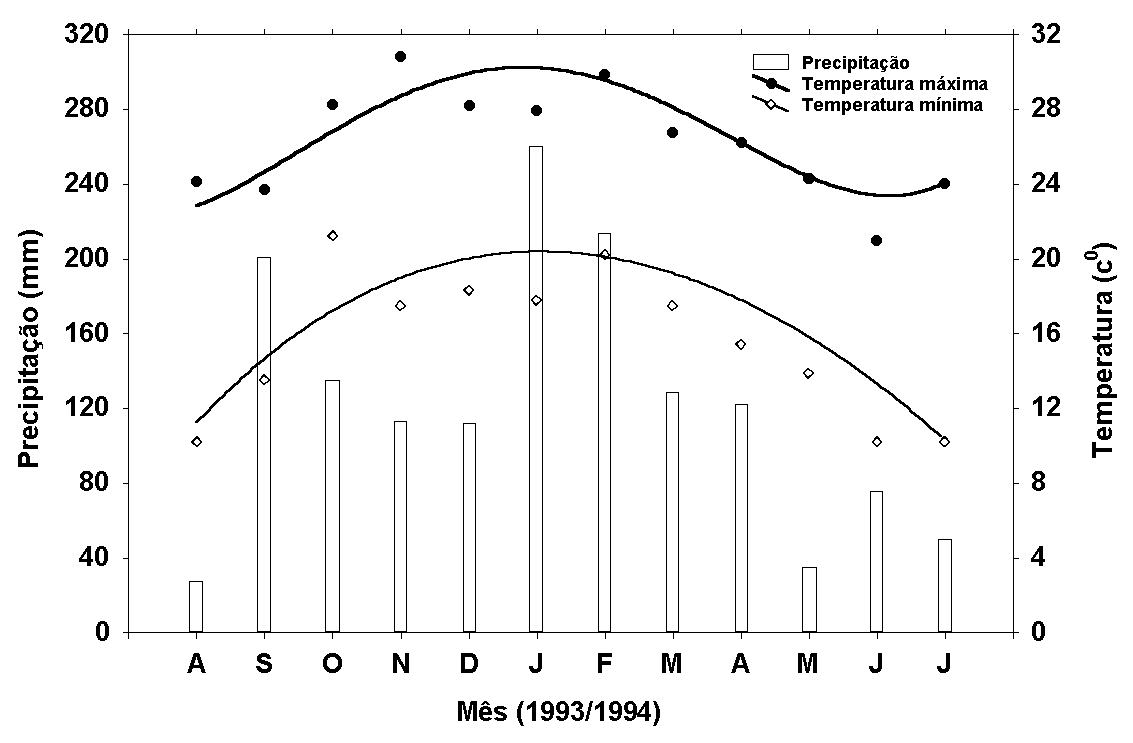
Figura 2:
Médias mensais das temperaturas máximas e mínimas (0C)
e a precipitação mensal (mm) obtidas no posto de observação
climatológica da Fazenda Holambra.
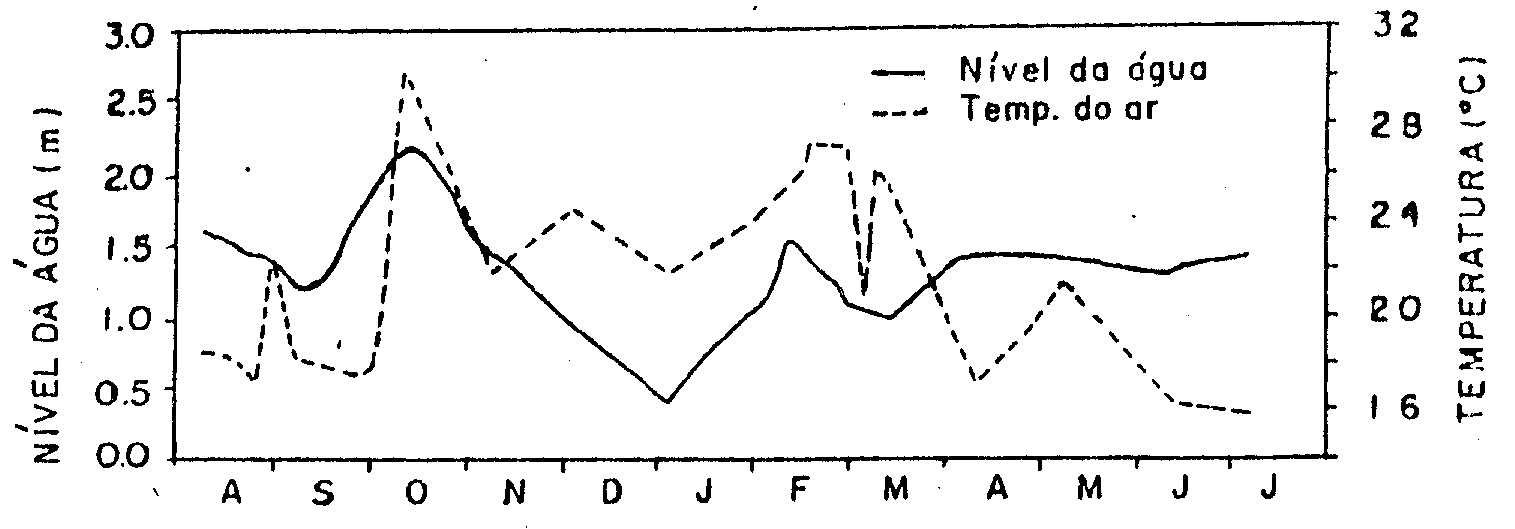
Figura 3: Altura
da régua (m) no interior do estande e temperatura do ar
(0C) no local de estudo.
A análise de agrupamento
(Figura 4), permitiu identificar dois períodos climatologicamente
distintos durante o ano de estudo. O primeiro compreendeu
os meses de menores temperaturas e precipitações (abril
a setembro) e o segundo período incluiu os meses de maiores
precipitações e temperaturas (de outubro a março).
A variação anual do
nível da água (cota) mostrou também uma oscilação bem marcada
(Figura 5). Entre o início de agosto e o final de setembro/93
ocorreu uma pequena diminuição do nível da água. Posteriormente,
em função da elevada precipitação ocorrida em meados de
setembro/93, houve uma rápida elevação, atingindo um pico
máximo no início de outubro/93. Seguiu-se um gradativo abaixamento
até janeiro/94 (período de menor nível da água). A partir
de fevereiro/94 ocorreu uma rápida elevação, devido principalmente
às intensas precipitações ocorridas em janeiro/94 e associadas
com as vazões mais baixas, propiciaram acúmulo de água no
reservatório. A partir do final de fevereiro e início de
março/94, o nível da água manteve-se relativamente estável
até julho/94.
Desta forma, foi observada
uma amplitude de mais de 2m na variação do nível da água.
Esse padrão de variação é em grande parte devido à operação
do reservatório, particularmente relacionado às alterações
de vazão.
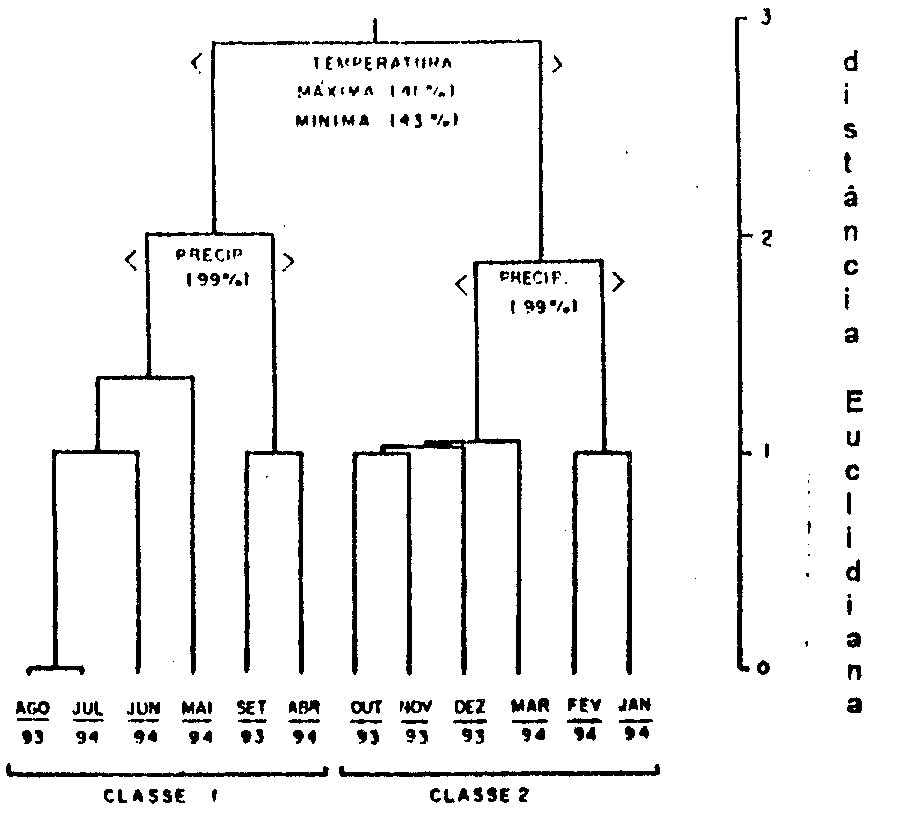
Figura 4:
Dendrograma baseado nos valores de temperaturas máximas
e mínimas e de precipitações, obtidos no posto de observação
climatológica da Fazenda Holambra.
A variação do nível
da água determinada no interior do estande de E. polystachya
(Figura 3), tem padrão de variação similar ao das cotas.
O modelo de superfície
Z=a+bX+cY+dX2+eXY+fY2,
onde Z= temperatura (0C), Y= profundidade (m),
X= mês e a, b, c, d, e
e f constantes, aplicado aos valores de temperatura
da água na superfície, meio e fundo mostrou-se estatisticamente
significativo (Figura 6). Para ambos os locais, observa-se
padrão semelhante na variação anual, com temperaturas mais
elevadas entre os meses de outubro/93 e março/94, coincidente
com o padrão observado para a temperatura do ar. A temperatura
do ar mostrou-se positivamente correlacionada com a temperatura
da água do rio (r=0,67, p<0,05) e do estande (r=0,65,
p<0,05). A temperatura da água do estande e do rio também
apresentaram-se correlacionadas (r=0,99, p<0,05).
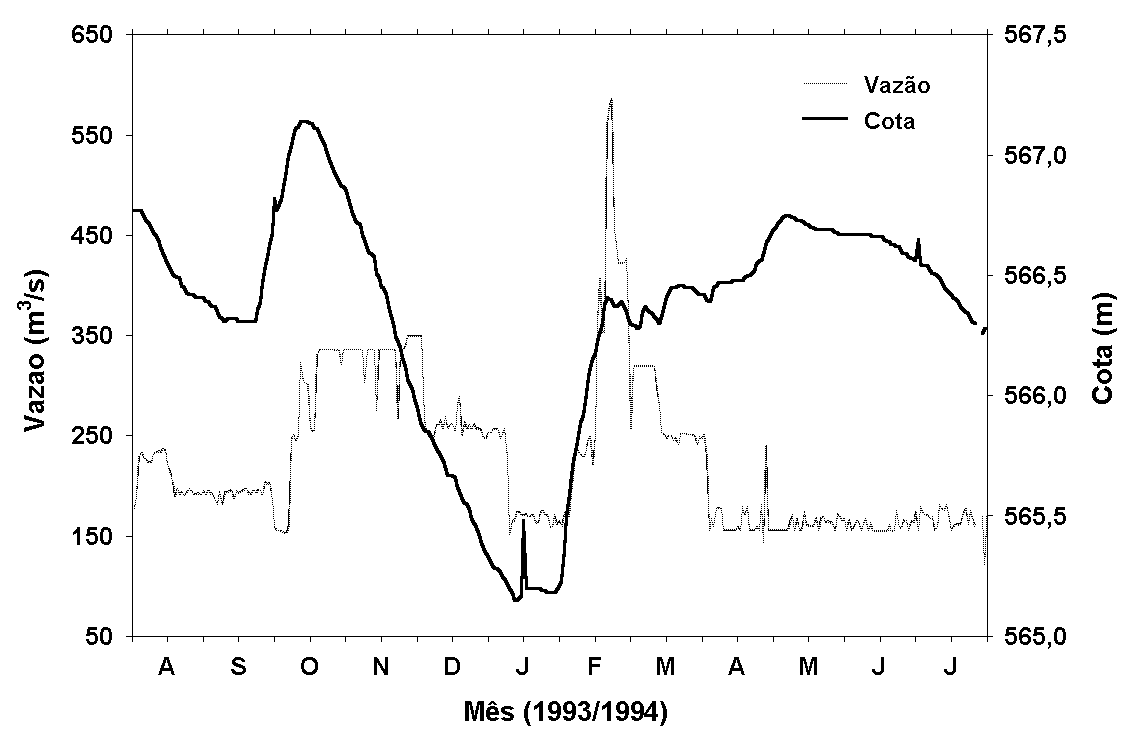
Figura 5:
Variação anual do nível da água (cota - m) e da vazão efluente
(m3/s) da represa de Jurumirim.
Em relação à
condutividade elétrica na coluna dágua, maiores valores
foram medidos nos meses de novembro/93 a março/94 (Figura
6). A menor e a maior condutividade elétrica determinada
na água do rio e do estande foram de 33,70 e 38,80m S/cm
e 87,00 e 78,40m S/cm, respectivamente. O seu padrão de
variação sazonal foi inverso aos observados para a precipitação
e para a concentração de MST (Figura 7a). Uma correlação
significativa entre os valores de condutividade elétrica
dos dois locais foi também encontrada (r=0,64, p<0,05).
No rio, a concentração
de MST apresentou padrão anual semelhante ao da precipitação.
A concentração de MST apresentou amplitude de variação de
2,76 a 47,87mg/l e 8,08 a 72,30mg/l, no rio e estande, respectivamente.
Maiores valores foram encontrados entre novembro/93 e março/94
(Figura 7a). Nenhuma relação significativa foi encontrada
entre as concentrações de MST na água do rio e na presente
na água do estande (r=-0,14, p<0,05).
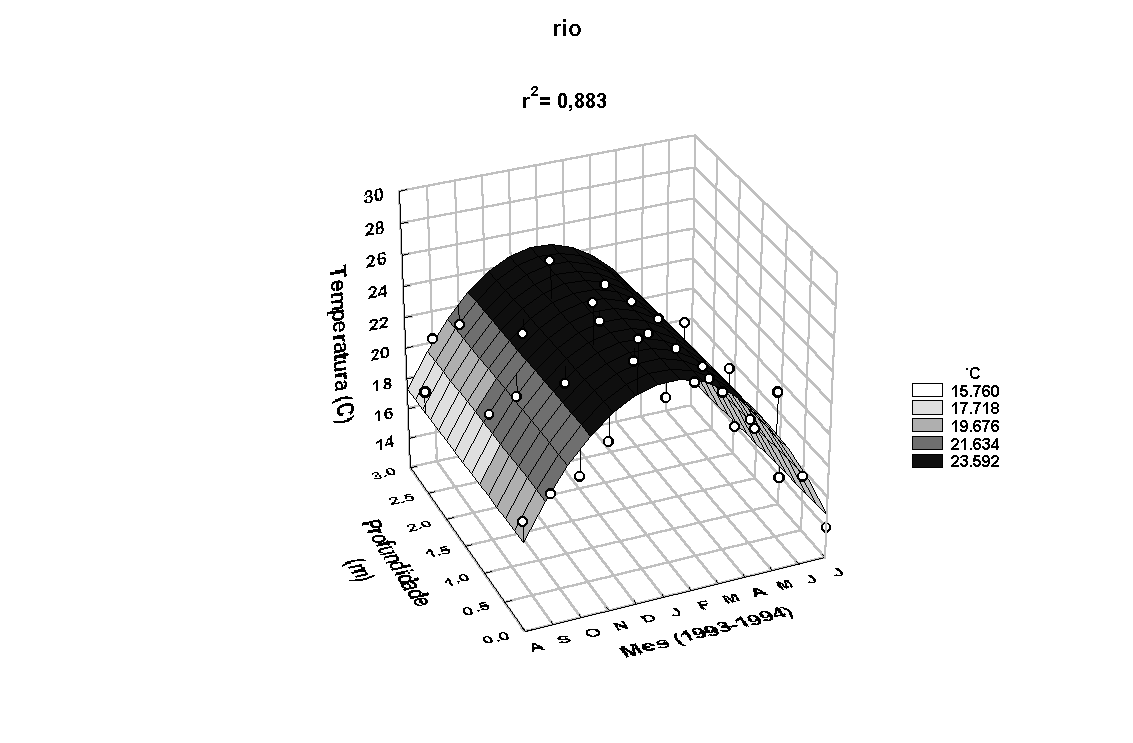 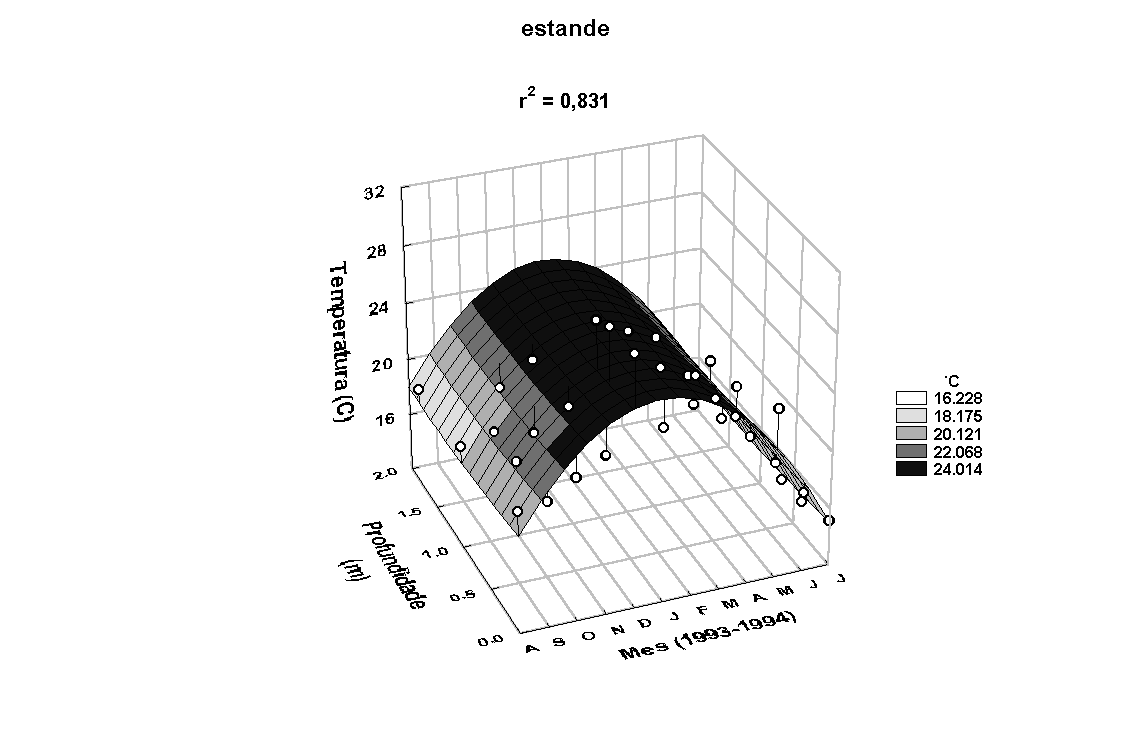 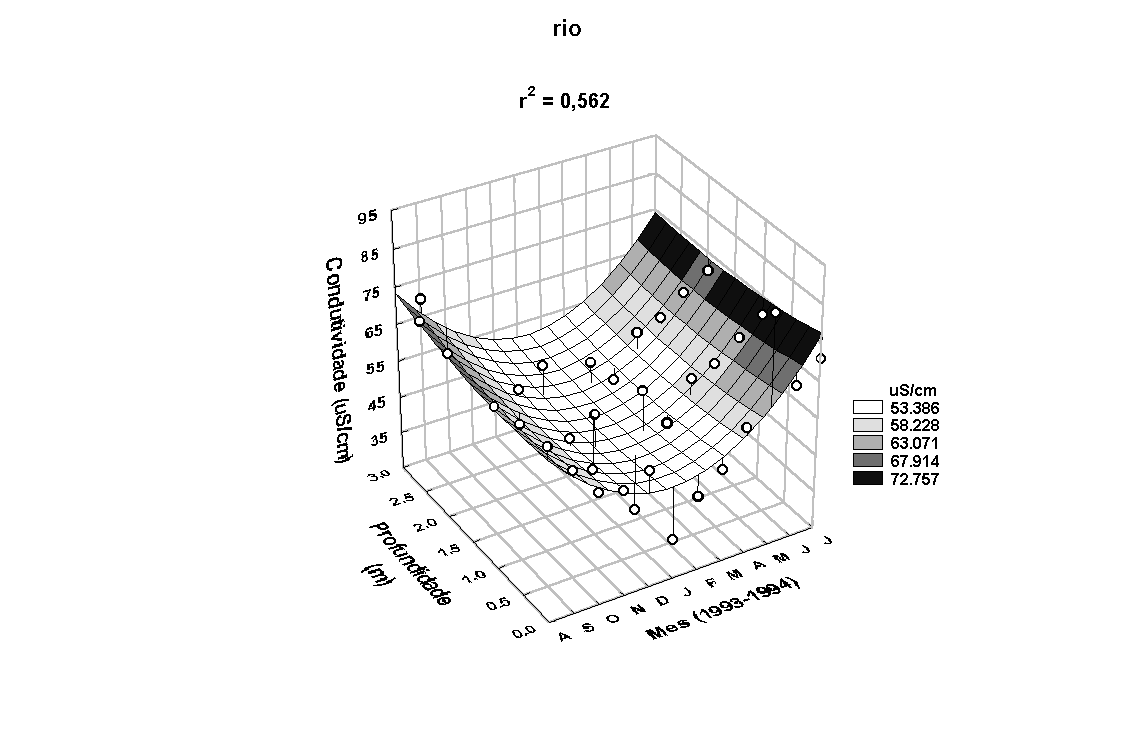 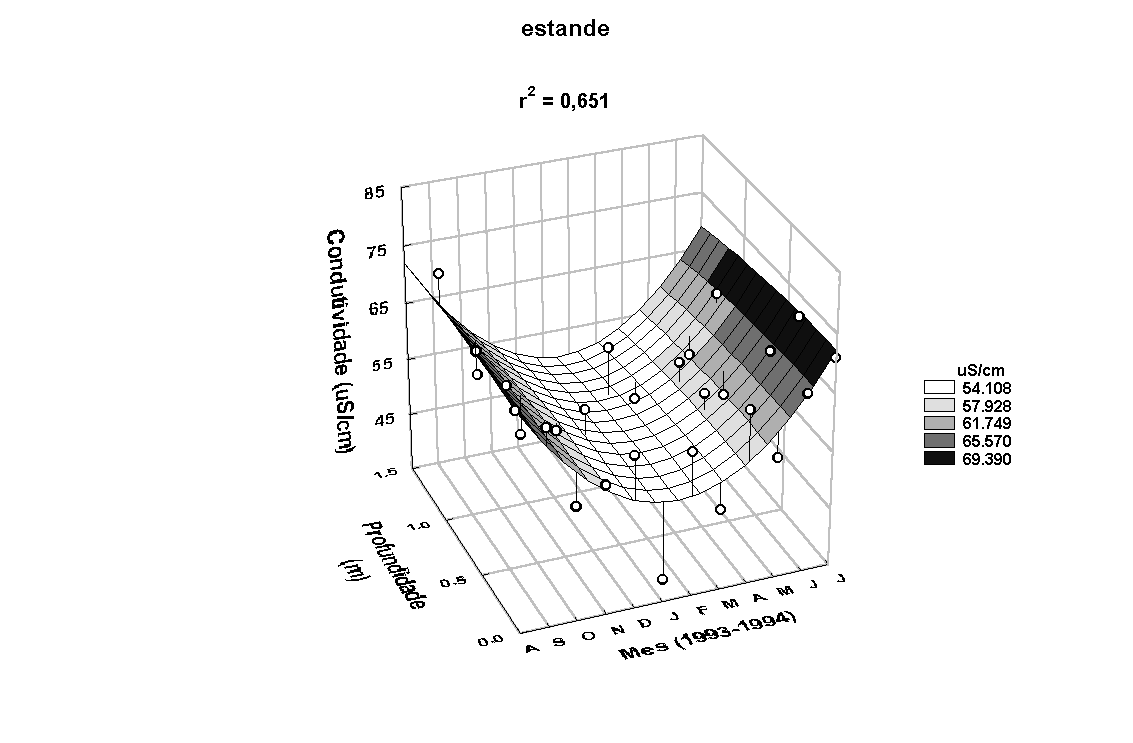
Figura 6: Variação anual da temperatura e da condutividade elétrica da água
no rio Paranapanema e no estande de E. polystachya.
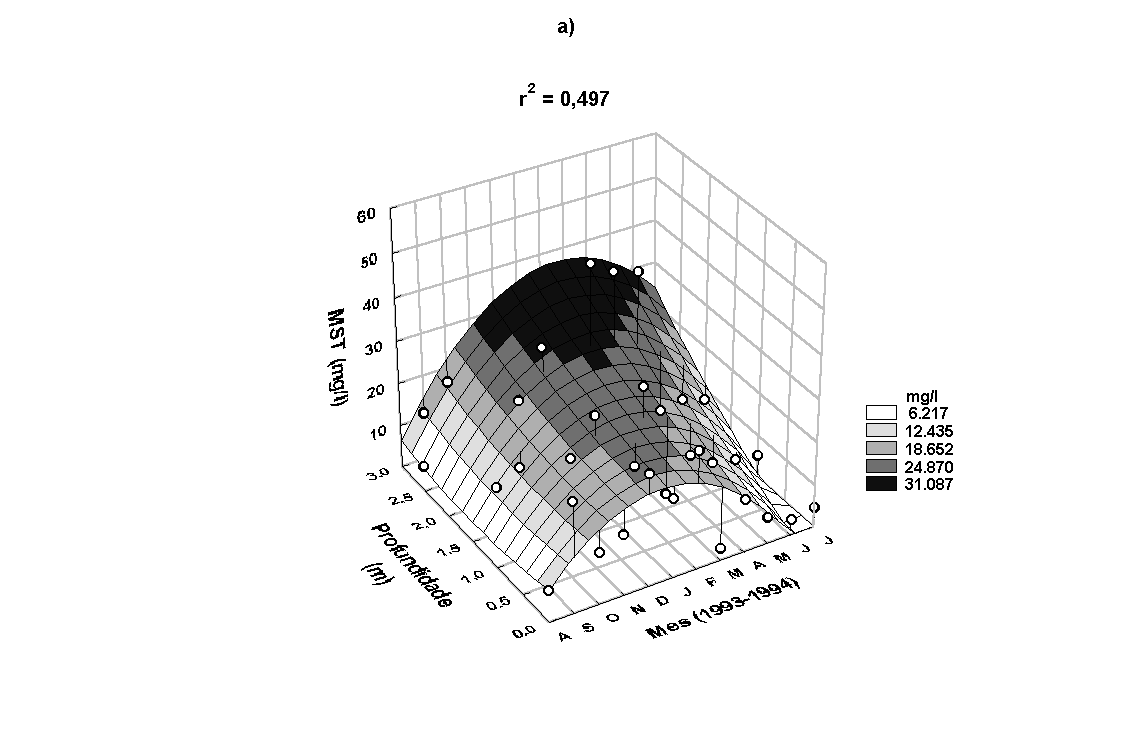 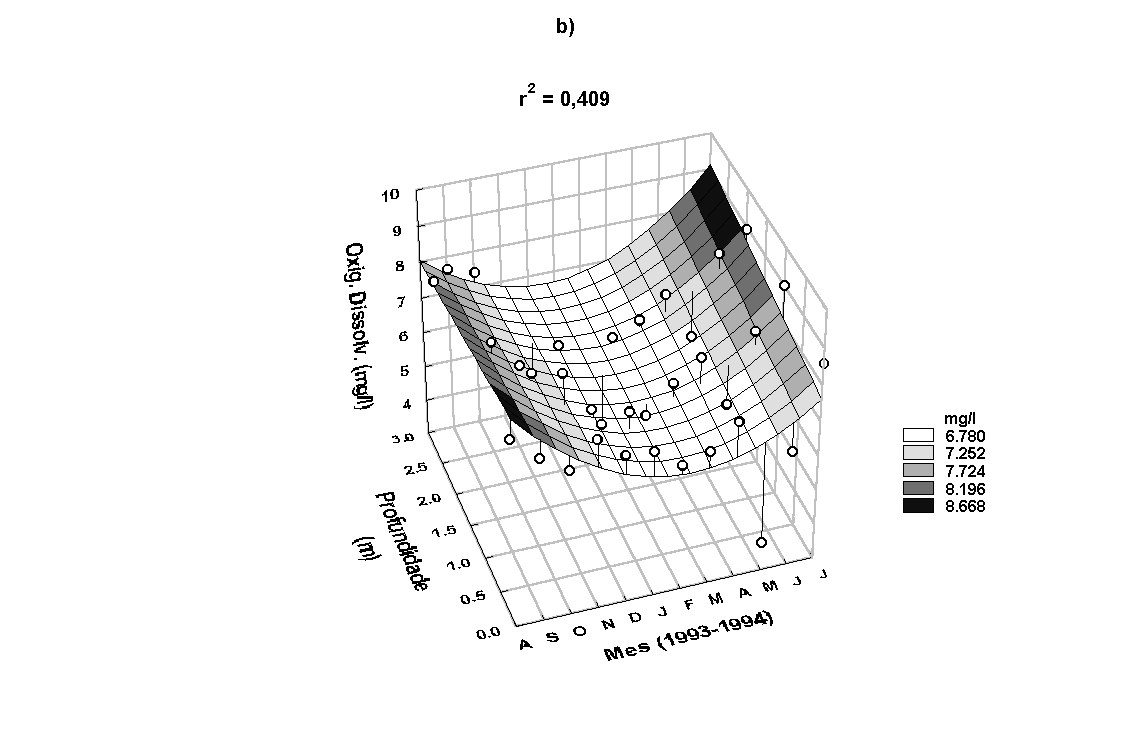
Figura 7:
Variação anual da concentração de MST (a) e de oxigênio
dissolvido na água do rio (b).
As menores concentrações
de oxigênio dissolvido na água do rio foram encontradas
entre os meses de novembro/93 a março/94 e as maiores de
agosto a outubro/93 e de abril a julho/94 (Figura 7b). O
padrão de variação anual apresentado pelo teor de oxigênio
dissolvido foi inverso ao da temperatura da água. O padrão
de variação sazonal da concentração de oxigênio dissolvido
determinado na água do estande é apresentado na Figura 8.
Também nenhuma correlação linear foi encontrada entre os
teores de oxigênio dissolvido
determinados na água do rio e no interior do estande (r=0,24,
p<0,05). Os teores de oxigênio dissolvido variaram de
3,78 a 8,97mg/l e 0,63 a 8,26mg/l, respectivamente para
a água do rio e do estande.
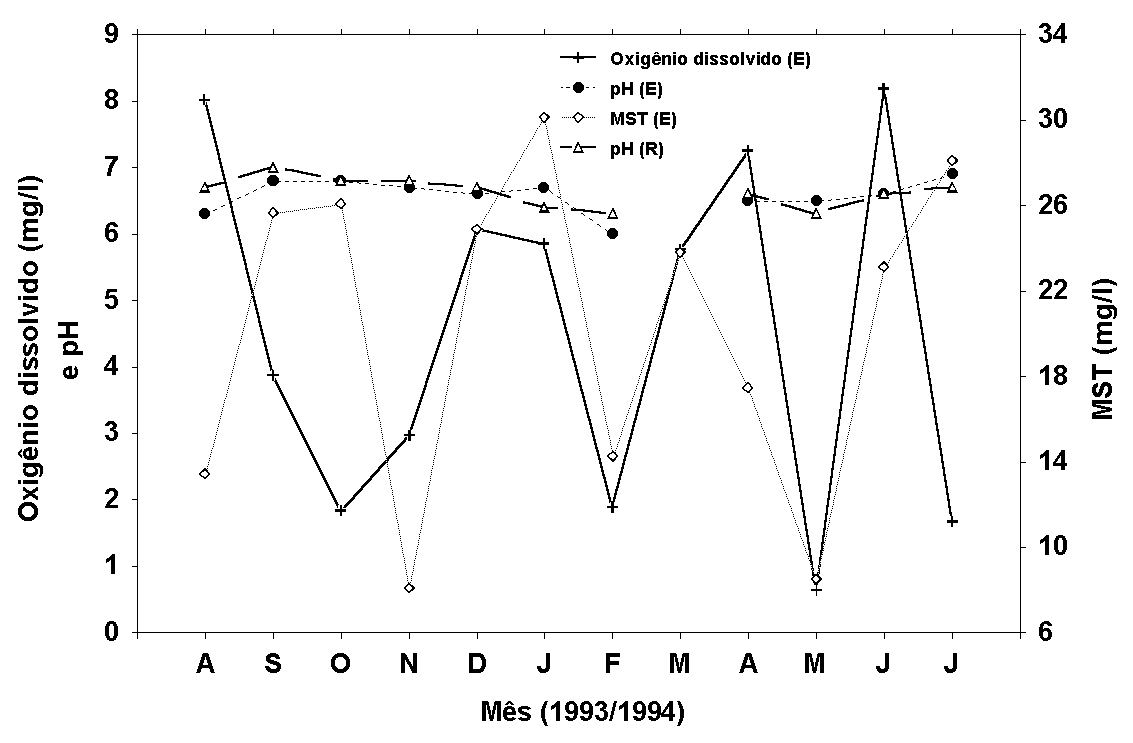
Figura 8:
Médias mensais (superfície, meio e fundo) do pH e valores
de superfície das concentrações de MST e de oxigênio dissolvido
(E - estande; R - rio).
O pH da água apresentou
pequena amplitude de variação ao longo do ano, de 6,1 a
7,1 no rio e de 6,0 a 7,1 no estande (Figura 8). Nenhuma
correlação significativa foi determinada entre o pH das
estações de coletas de água (r=-0,14, p<0,05).
Em ambas as estações
de coletas, as concentrações de nitrito apresentaram valores
baixos e nenhuma correlação foi encontrada entre elas (Figura
9, r=-0,08, p<0,05). Os valores de nitrito variaram de
abaixo do limite de detecção do método a 6,84m g/l no rio
e 0,91 a 3,91m g/l no estande.
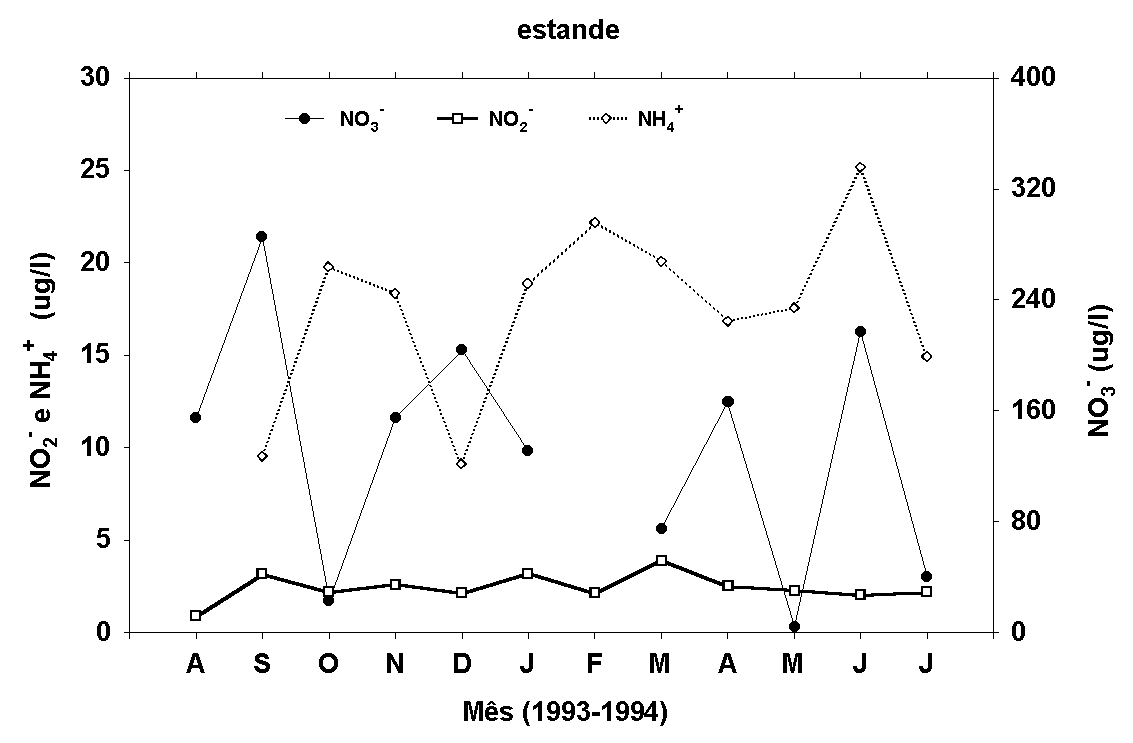 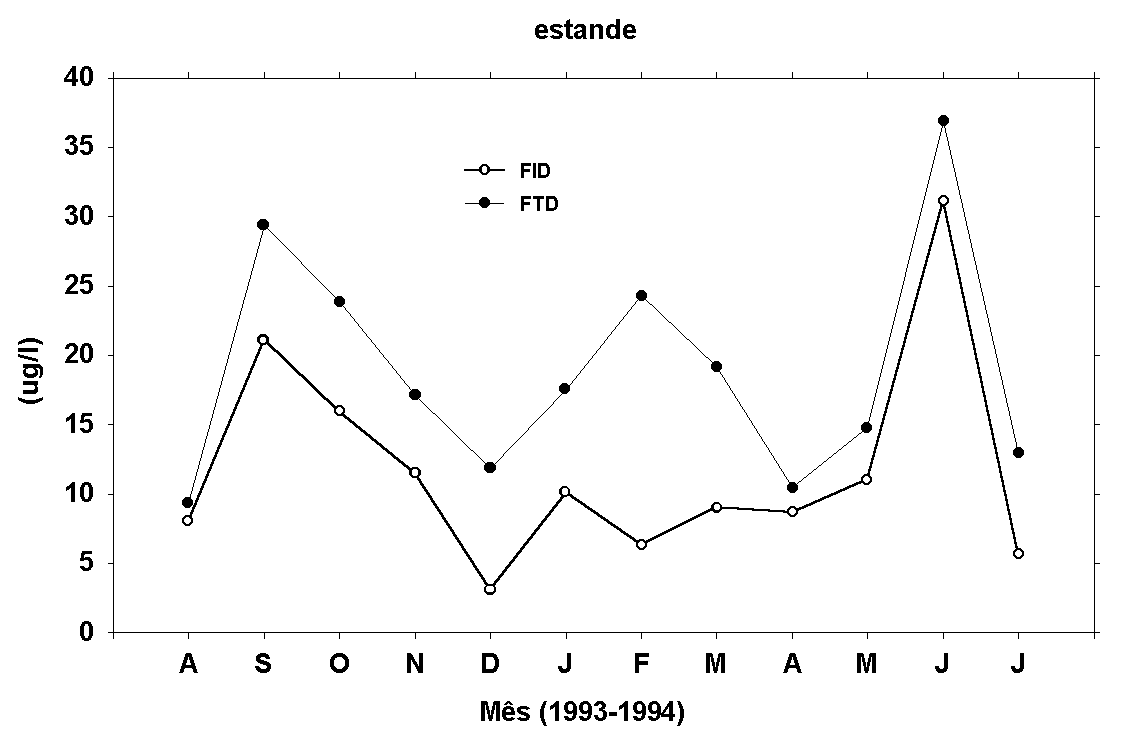
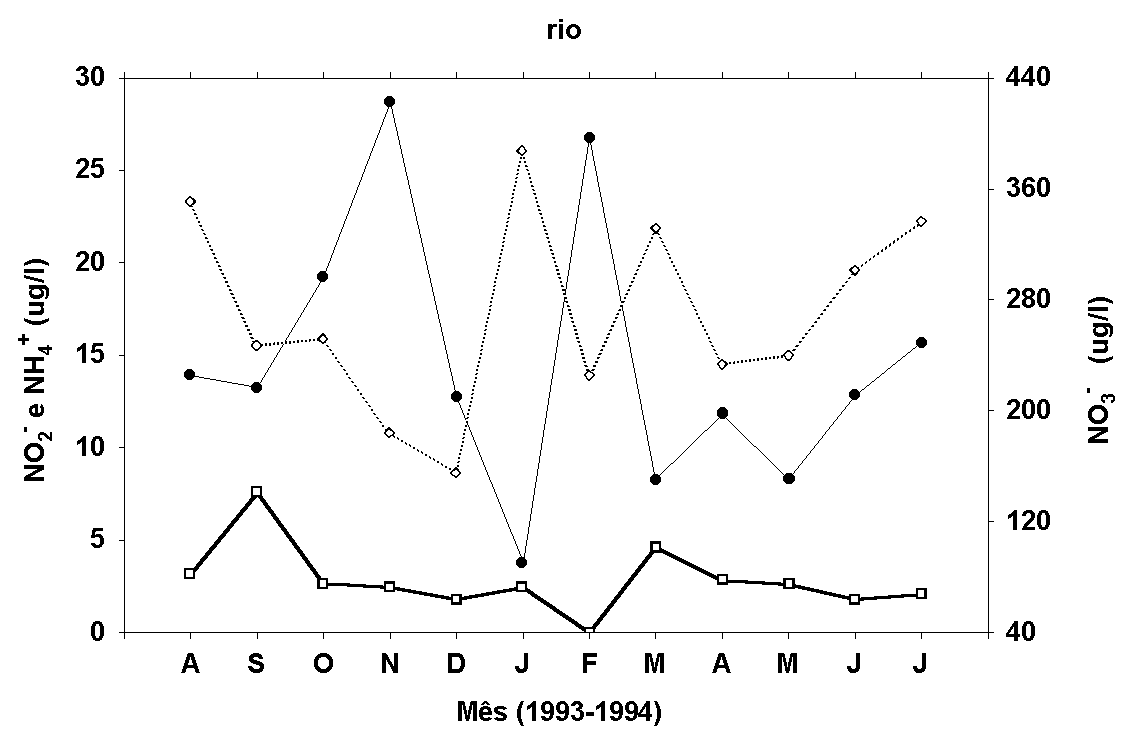 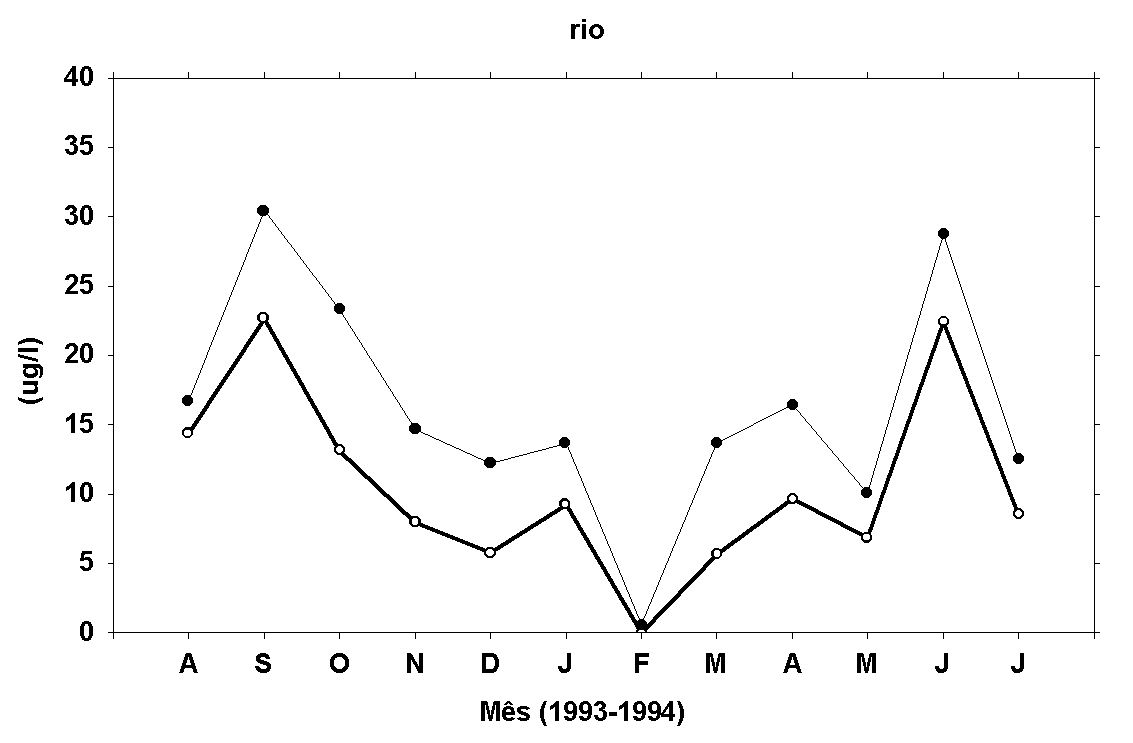
Figura 9:
Médias mensais (superfície, meio e fundo) dos teores de
nitrito, amônia, FID e FTD na água do rio e do estande.
Para o teor de nitrato (estande) são apresentados os valores
da água superficial.
Não foi verificada correlação
linear entre as concentrações de nitrato e de amônia determinadas
na água do rio e do estande (NO3- - r=0,16; NH4+
- r=-0,04, p<0,05), mas um padrão de variação inverso
entre os valores de amônia e de nitrato é sugerido (Fig.
9). Os teores de nitrato variaram de 12,95 a 577,31m g/l
no rio e abaixo do limite de detecção do método a 330,31m
g/l no estande. Os de amônio variaram de 2,34 a 67,45m g/l
no rio e 5,10 a 30,01m g/l no estande.
Os teores de fosfato,
em geral, apresentam menores valores no final de 93 e início
de 94, com exceção de FTD na água do rio, que apresentou
um pico em fevereiro/94 (Figura 9). As concentrações de
FID determinadas no rio e no estande estão correlacionadas
entre si (r=0,62, p<0,05). Em relação aos teores de FTD,
determinados na água dos dois locais, nenhuma correlação
linear significativa foi encontrada (r=0,43, p<0,05).
Os teores de FTD apresentaram a seguinte amplitude de variação:
abaixo do limite de detecção do método a 51,45m g/l no rio
e 9,34 a 57,21m g/l no estande. Os teores de FIT variaram
de a abaixo do limite de detecção do método tanto no rio
e no estande a 44,17m g/l (rio) e 51,16m g/l (estande).
Tabela 1: Valores
médios das variáveis físicas e químicas para superfície
(S), meio (M) e fundo (F) e resultados do teste estatístico
multivariado (p<0,05).
| variável |
S |
M |
F |
Fcal |
Fcrit |
conclusão |
|
|
|
estande |
|
|
|
| oxigênio
(mg/l) |
4,01 |
5,10 |
5,50 |
4,76 |
4,46 |
S¹(M=F) |
| pH |
6,6 |
6,5 |
6,5 |
0,14 |
4,46 |
S=M=F |
cond.
elétrica (mS/cm) |
62,05 |
61,50 |
60,57 |
3,53 |
4,26 |
S=M=F |
| MST
(mg/l) |
18,74 |
32,55 |
33,70 |
5,81 |
5,79 |
S¹(M=F) |
NO3-
(mg/l) |
101,73 |
188,80 |
200,46 |
6,24 |
5,14 |
S¹(M=F) |
NO2-
(mg/l) |
2,04 |
2,25 |
2,55 |
3,19 |
5,14 |
S=M=F |
NH4+
(mg/l) |
18,22 |
18,49 |
18,19 |
0,01 |
5,14 |
S=M=F |
FID
(mg/l) |
10,04 |
14,98 |
11,69 |
0,36 |
5,14 |
S=M=F |
FTD
(mg/l) |
18,61 |
21,05 |
17,59 |
0,37 |
5,14 |
S=M=F |
| temperatura
(0C) |
22,07 |
21,57 |
21,42 |
4,47 |
4,26 |
S¹(M=F) |
|
|
|
rio |
|
|
|
| oxigênio
(mg/l) |
7,05 |
7,37 |
7,15 |
1,76 |
4,10 |
S=M=F |
| pH |
6,6 |
6,6 |
6,7 |
1,35 |
4,26 |
S=M=F |
cond.
elétrica (mS/cm) |
59,49 |
57,92 |
58,24 |
0,17 |
4,10 |
S=M=F |
| MST
(mg/l) |
13,40 |
13,28 |
23,01 |
2,81 |
4,10 |
S=M=F |
NO3-
(mg/l) |
236,14 |
246,51 |
219,74 |
0,53 |
4,10 |
S=M=F |
NO2-
(mg/l) |
3,11 |
2,62 |
2,63 |
1,56 |
4,10 |
S=M=F |
NH4+
(mg/l) |
14,54 |
14,21 |
23,02 |
1,33 |
4,10 |
S=M=F |
FID
(mg/l) |
11,13 |
8,56 |
12,27 |
1,15 |
4,10 |
S=M=F |
FTD
(mg/l) |
17,02 |
14,06 |
17,61 |
0,98 |
4,10 |
S=M=F |
| temperatura
(0C) |
21,89 |
21,56 |
21,49 |
2,51 |
4,10 |
S=M=F |
A análise multivariada
indica que os valores das variáveis físicas e químicas determinadas
em várias profundidades no rio não diferem do ponto de vista
estatístico, sugerindo que não há gradientes verticais entre
as variáveis medidas. O mesmo não foi observado para o estande.
A comparação revelou que houve menores teores de oxigênio
dissolvido, MST e nitrato e maior temperatura na camada
superficial de água do estande, quando comparado com os
valores determinados nas camadas mais profundas. Para estas
variáveis um padrão de estratificação foi encontrado (Tabela
1).
Tabela 2: Valores
médios dos parâmetros físicos e químicos ao longo do ano
para rio (R), baía (B) e estande (E) e resultados do teste
estatístico multivariado (p<0,05).
| variável |
Baía(1) |
Estande |
Rio |
Fcal |
Fcrit |
conclusão |
| oxigênio
(mg/l) |
6,86 |
4,49 |
7,19 |
6,78 |
4,10 |
E¹(R=B) |
| pH |
6,5 |
6,7 |
6,6 |
0,52 |
4,10 |
E=R=B |
| cond.
elétrica (mS/cm) |
55,12 |
58,75 |
58,55 |
1,86 |
4,10 |
E=R=B |
| MST
(mg/l) |
7,12 |
18,31 |
16,18 |
22,15 |
4,10 |
B¹(E=R) |
| NO3-
(mg/l) |
155,06 |
121,54 |
234,54 |
3,79 |
4,10 |
E=R=B |
| NO2-
(mg/l) |
1,90 |
2,44 |
2,59 |
1,34 |
4,10 |
E=R=B |
| NH4+
(mg/l) |
14,69 |
17,48 |
17,13 |
0,49 |
4,10 |
E=R=B |
| FID
(mg/l) |
5,40 |
11,82 |
10,59 |
4,09 |
4,10 |
E=R=B |
| FTD
(mg/l) |
13,46 |
18,97 |
16,18 |
1,51 |
4,10 |
E=R=B |
| temperatura
(0C) |
22,25 |
22,22 |
21,64 |
6,01 |
4,10 |
R¹(E=B) |
1 - Dados obtidos de MOSCHINI-CARLOS19.
Também pôde ser verificado
através da análise multivariada diferenças estatisticamente
significativas (p<0,05) entre os teores de oxigênio dissolvido
na água do rio, do estande e da baía (Tabela 2), com valores
maiores no rio e na baía. Com relação ao MST, menores concentrações
são encontradas na baía, quando comparadas com aquelas do
estande e do rio. A comparação dos valores revela que no
rio há tendência de menor temperatura da água do que no
estande e na baía.
DISCUSSÃO
De acordo com Nimer
(20), a região do Alto Paranapanema apresenta temperatura
média anual quase sempre inferior a 220C. No
verão a temperatura média é superior a 220C.
Os dados climatológicos provenientes do posto meteorológico
da Fazenda Holambra e as medições efetuadas no local de
estudo corroboram o padrão geral descrito por Nimer (20).
A temperatura média
anual da água do rio Paranapanema foi de 21,60C,
com uma amplitude de variação ao longo do ano de 14,70C.
Drago (6) determinou para o rio Paraná (PR, Brasil), maiores
amplitudes de variação entre superfície e fundo (17,10C
e 15,60C, respectivamente). Thomaz et al. (28)
registraram amplitudes de variação, ao longo do ano, de
11,6 e 13,30C, respectivamente para os rios Paraná
e Ivinheima.
A variação anual da
temperatura da água do rio Paraná (2, 28) e do Arroyo Toledo-Carrasco
(Uruguai) (1), apresentou padrão de variação semelhante
ao do rio Paranapanema, com temperaturas máximas entre dezembro
e março e mínimas entre junho e agosto. O elevado coeficiente
de correlação encontrado reflete a dependência da temperatura
da água em relação à temperatura do ar, como assinalou Drago
(6). Segundo Hynes (13), para grandes rios e a uma considerável
distância de sua cabeceira, a temperatura da água é usualmente
próxima à média mensal da temperatura do ar. Cole (3) também
afirma que a temperatura da água apresenta um padrão de
variação anual semelhante ao da temperatura do ar. Desta
forma, como apontado por Schäfer (22), a temperatura, considerada
um dos principais fatores climáticos, tem papel importante
nas condições hidrológicas e ecológicas de um rio.
Drago (6) não observou
diferenças significativas entre os perfis de temperatura
da água do meio do canal e de ambas as margens do rio Paraná.
Atribuiu essa homogeneidade ao processo de mistura originado
pelo fluxo turbulento, que raramente propicia estratificação
térmica vertical da água nos grandes rios. Embora o rio
Paranapanema seja um curso de água com dimensões menores,
a homogeneidade sugerida na análise multivariada no canal
principal do rio pode ser explicada através da movimentação
da massa de água propiciando sua mistura.
A relação inversa entre
a variação anual da concentração de oxigênio dissolvido
e a temperatura da água, é decorrente do fato de que a solubilização
do oxigênio na água é função inversa à temperatura. Desta
forma, nos meses de temperaturas mais elevadas, a menor
solubilização desse gás acarreta teores mais baixos na água.
O aumento da temperatura também acarreta elevação nas taxas
de decomposição (14, 25), como conseqüência, pode ocorrer
a diminuição dos teores de oxigênio no meio. As concentrações
de oxigênio nos rios, segundo Schäfer (22), variam em função
das condições climáticas regionais e de sua localização
(em altitude ou na planície). Dessa forma, a marcada variação
anual do teor de oxigênio dissolvido observada, pode ser
explicada também em função das características meândricas
e hidrodinâmicas do rio Paranapanema, particularmente a
montante do local de estudo. Apesar de acarretar mistura
na massa d'água, provavelmente a turbulência não causa oxigenação,
e consequentemente alterações substanciais nos teores de
oxigênio dissolvido na água. Além disso, a montante do local
de estudo, possivelmente não há fontes pontuais (esporádicas
ou sazonais) significativas de matéria orgânica, que possam
propiciar uma diminuição dos teores de oxigênio na água
através do consumo pela decomposição.
O aumento na concentração
de MST observado no período de precipitações mais intensas,
deve ser atribuído provavelmente à maior participação proporcional
da fração inorgânica do material particulado transportado
pelo rio. Esse material alóctone, incluindo íons, deveria
propiciar aumento na condutividade elétrica. A maior entrada
de água proveniente da bacia de drenagem resulta numa diminuição
proporcional das concentrações de íons dissolvidos e, conseqüentemente,
na condutividade elétrica, caracterizando portanto um efeito
de diluição. Henry (dados não publicados),
trabalhando em Campina
do Monte Alegre, a montante do local deste estudo, demonstrou
através do desenvolvimento de modelo, a diminuição da condutividade
elétrica com o aumento da vazão, corroborando o efeito de
diluição como hipótese explicativa. Thomaz et al. (27) também
atribuiu às menores concentrações de nutrientes observadas
no rio Baía (MS, Brasil) no período de cheia à diluição
decorrente da subida das águas do rio Paraná. As menores
concentrações das frações de fósforo observadas na água
do rio e no estande no período de maior precipitação também
atestam o efeito de diluição.
Com relação ao padrão
de variação alternado dos teores de nitrato e de amônia
sugerido, processos relacionados à ciclagem do nitrogênio,
particularmente à nitrificação, devem ser cogitados na explicação
dessas oscilações.
Thomaz et al. (28) não
observaram padrão tão marcante na variação do nível da água
do rio Paraná e Ivinheima, quando comparado com o do rio
Paranapanema. Segundo esses autores, o controle da vazão
do reservatório de Itaipu se faz sentir sobre os níveis
fluviométricos do rio Paraná, especialmente durante o período
de seca. Na zona de desembocadura do rio Paranapanema, o
padrão sazonal observado no nível da água é decorrente principalmente
da operação do reservatório, com alterações de vazão em
função da demanda. Uma defasagem de cerca de um mês entre
o período de maiores precipitações e a elevação da cota
do reservatório foi também constatada.
Em Campina do Monte
Alegre (rio Paranapanema, a montante de sua zona de desembocadura),
Henry (9) determinou através de coletas bimensais de maio/88
a março/89, uma variação nas concentrações de nitrato de
74,2 a 137,0mg/l; de FTD de 2,57 a 59,0mg/l e de MST de
14,5 a 67,3mg/l. No presente estudo, as concentrações de
FTD e de MST foram semelhantes às obtidas por Henry (9).
Em relação ao nitrato, concentrações mais altas e com maior
variabilidade foram encontradas. Estas diferenças podem
ser atribuídas à maior intensidade das amostragens efetuadas
neste estudo e a influxos das zonas marginais, como as lagoas.
Segundo Howard-Williams
& Lenton (12), na zona litoral de lagos com grande cobertura
de plantas aquáticas, a desoxigenação da água é devida principalmente
a três fatores: (a) reduzida movimentação da massa de água
em função da presença da cobertura vegetal interferindo
na oxigenação, (b) baixas taxas de fotossíntese fitoplanctônica
na região de macrófitas aquáticas e, conseqüentemente, baixa
produção de oxigênio, em função da menor intensidade luminosa
e (c) rápido consumo do oxigênio produzido pelas plantas
por organismos presentes no interior dos estandes. Além
desses fatores, a temperatura mais elevada observada na
superfície da água no estande reduz a solubilização desse
gás e, consequentemente, a sua concentração na água. Kunii
et al. (16) observaram período de considerável diferença
na concentração de oxigênio dissolvido entre a superfície
e o meio com o fundo em Ojaga-ike (Japão), uma lagoa rasa
e eutrófica. Segundo os autores, a maior concentração de
oxigênio dissolvido observada na superfície dessa lagoa
pode ser devida à sua liberação, durante a atividade fotossintética
das macrófitas aquáticas na camada superior, ou ainda devido
ao seu maior consumo pela decomposição que ocorre no fundo,
acarretando diferenças elevadas entre a superfície e o fundo.
Provavelmente, os fatores assinalados anteriormente, atuando
em conjunto, devem ser os responsáveis pelo padrão de variação
anual da concentração de oxigênio dissolvido no estande
de E. polystachya. Esses fatores também devem ser
os responsáveis pelo gradiente constatado no estande entre
superfície e meio e fundo. Os menores teores de oxigênio
dissolvido determinados na água do estande de E. polystachya,
quando
comparados com os teores
determinados na água do rio e da baía, podem ser explicados
por processos mais intensos de decomposição no estande do
que no rio, ocasionando maior consumo de oxigênio e, consequentemente,
diminuição dos seus teores na água. Assim, os teores de
oxigênio dissolvido determinados na água do rio refletem
principalmente influências externas, relacionada ao padrão
sazonal de variação da temperatura do ar. As concentrações
determinadas na água presente no interior do estande de
E. polystachya refletem principalmente processos
internos, como produção e decomposição.
As diferenças significativas
entre as temperaturas e as concentrações de oxigênio dissolvido
encontradas na água do rio e no interior do estande, bem
como o gradiente térmico observado no estande, pode ser
explicado pela presença do estande de E. polystachya.
Este atua como barreira física à passagem da água e à ação
do vento. Assim, haveria menor renovação da água, com seu
tempo de residência aumentado e a mistura da massa dágua
pela ação do vento seria reduzida. A falta de correlação
entre as variáveis físicas e químicas da água do rio e do
estande, também ressalta as diferenças entre as massas de
água.
As diferenças observadas
entre as concentrações de oxigênio dissolvido, de MST e
da temperatura na água do rio, do estande e da baía, podem
estar relacionadas à variação do nível da água (2, 25, 26,
28). Como a baía estudada é rasa e, em períodos de águas
baixas, encontra-se mais isolada do rio (19), estas características
sugerem que processos internos, como a circulação da massa
dágua pela ação do vento, também devem ser responsáveis
pelas diferenças apontadas. Em decorrência das baixas profundidade
e velocidade da corrente, além da presença de períodos com
estratificação térmica, a baía estudada pode ser considerada
como um ambiente intermediário entre lêntico e lótico de
natureza polimítica (19).
AGRADECIMENTOS
À FAPESP (Projeto Temático:
proc. 91/0612-5) pelo suporte financeiro e pela bolsa de
estudos (proc. 92/2886-8) e aos Srs. H.A. Rodrigues e M.F.P.
de Oliveira pelo auxílio nos trabalhos de campo e de laboratório.
ABSTRACT
The influence of
the macrophyte Echinochloa polystachya (H.B.K.) Hitchcock
on the physical and chemical characteristics of water in
the mouth zone of Paranapanema River into the Jurumirim
Reservoir - SP. In the mouth zone of Paranapanema River
into the Jurumirim Reservoir (SP, Brazil), the emergent
macrophyte E. polystachya dominate the vegetal physiognomy.
This plant is rooted into the marginal area of the water
body and stems towards the pelagic region. The seasonal
patterns of physical
and chemical variables of water in the stand of this grass
and in the main river channel were studied. The annual variation
of the water temperature and dissolved oxygen concentrations
is due to the modifications on the air temperature. The
changes on the concentrations of suspended matter and on
the electric conductivity were associated to the precipitation
regime during the year. An homogeneity on the physical and
chemical factors in the river was observed and was due to
the movement of the water mass. In the stand, the surface
layer presented different characteristics when compared
with the intermediary and deep zones. A comparison between
the values of temperature and dissolved oxygen concentrations
in the stand water and in the river showed that the differences
were statistically significant. These can be attributed
to the presence of E. polystachya, that acts like
a physical barrier on the water and to the wind action.
Key-words: macrophyte,
Echinochloa polystachya, dam, Paranapanema River,
hydrological cycle, physical and chemical characteristics
REFERÊNCIAS
- Arocena, R.; Chalar, G.; de Leon,
R.; Pintos, W. Evolucion anual de algunos parametros físico-químicos
del arroyo Toledo-Carrasco (Uruguay). Acta Limnol. Brasil.,
4:225-237, 1992.
- Camargo, A.F.M.; Esteves, F.A. Influence
of water level variation on fertilization of an oxbow
lake of rio Mogi-Guaçu, State of São Paulo, Brazil. Hydrobiologia,
299:185-193, 1995.
- Cole, G. Textbook of Limnology, 2
ed. The C.V. Mosby Company. Missouri, 1979, 425 p.
- Curi, P.R. Análise de agrupamento:
métodos seqüênciais, aglomerativos e hierárquicos. Ciênc.
e Cult., 35(10):1416-1429, 1983.
- Dale, H.K.; Gillespie, T.J. Diurnal
temperature gradients in shallow water produced by populations
of artificial aquatic macrophytes. J. Can. Bot., 56(9):1099-1106,
1978.
- Drago, E.C. Estudios limnologicos
en una seccion transversal del tramo medio del rio Parana.
VI Temperatura del agua. Rev. Asoc. Cienc. Nat. Litoral,
15:79-92, 1984.
- Esteves, F.A. Fundamentos de Limnologia,
Interciência/FINEP. Rio de Janeiro, 1988, 575 p.
- Golterman, H.L.; Clymo, R.S.; Ohnstad,
M.A.M. Methods for physical and chemical analysis of freshwaters.
20 ed. Blackwell Scientific Publications. Oxford, 1978,
213 p. (I.B.P. Handbook, 8).
- Henry, R. A exportação de nutrientes
e material em suspensão em alguns cursos de água na bacia
do Alto Paranapanema (Estado de São Paulo). Acta Limnol.
Brasil., 4:67-79, 1992.
- Henry, R. Amônia ou fosfato como
agente estimulador do crescimento do fitoplâncton na represa
de Jurumirim (rio Paranapanema, SP)?. Rev. Brasil. Biol.,
50(4):883-892, 1990.
- Henry, R. Primary production by phytoplankton
and its controlling factors in Jurumirim Reservoir (São
Paulo, Brasil). Rev. Brasil. Biol. 53(4):489-499, 1993.
- Howard-Williams, C.; Lenton, G.M.
The role of the littoral zone in functioning of a shallow
tropical lake ecosystem. Freshwat. Biol., 5:445-459, 1975.
- Hynes, H.B.N. The ecology of running
waters. University of Toronto Press. 1970, 555p.
- Junk, W.J. Áreas inundáveis: Um desafio
para Limnologia. Acta Amazonica,. 10(4):775-795, 1980.
- Koroleff, F. Determination of nutrients.
In: Grasshoff, K. (ed.) Methods of seawater analysis.
Verlag Chemie Weinhein, 1976, p. 117-181.
- Kunii, H.; Maeda, K.; Sastroutomo,
S.S.; Ikusima, I. Seasonal changes of water quality in
Ojaga-ike (pond) dominated by aquatic macrophytes in summer,
in Chiba, Japan. Bull. Mar. Lab., 12:31-40, 1980.
- Mackereth, J.F.H; Heron, J.; Talling,
J.F. Water analysis: some revised methods for limnologists.
Freshwater Biological Association, n0 36, 121
p. 1978.
- Morrison, D.F. Multivariate statistical
methods. McGraw-Hill, 415p., 1976.
- Moschini-Carlos, V. Estrutura e dinâmica
da comunidade perifítica em substrato natural e artificial
na represa de Jurumirim (zona de desembocadura do rio
Paranapanema, SP - Brasil), São Carlos, 1996, 171 p. (Tese,
Universidade Federal de São Carlos)
- Nimer, E. Climatologia do Brasil,
IBGE. Rio de Janeiro, 1979, 422p.
- Pompêo, M.L.M. Ecologia de Echinochloa
polystachya (H.B.K.)
- Hitchcock na represa de Jurumirim
(zona de desembocadura do rio Paranapanema-SP). São Carlos,
1996, 150 p. (Tese, Universidade de São Paulo)
- Schafer, A. Fundamentos de ecologia
e biogeografia das águas continentais. Editora da Universidade,
Porto Alegre, 1985. 532p.
- Strickland, J.D.; Parsons, T.R. A
manual of seawater analysis. Bull. Fihs. Res. Bul. Can.,
125:1-185, 1960.
- Teixeira, C.; Tundisi, J.G.; Kutner,
M.B. Plankton studies in a mangrove. II. The standing-stock
and some ecological factors. Bol. Inst. Oceanogr., 24:23-41,
1965.
- Thomas, S.M.; Lansac Toha, F.A.;
Roberto, M.C.; Esteves, F.A. Lima, A.F.; Seasonal of some
limnological factors of lagoa do Guaraná, a várzea lake
of the High rio Paraná, State of Mato Grosso do Sul, Brazil,
Rev. Hydrobiol. trop., 25(4):269-276, 1992a.
- Thomas, S.M.; Roberto, M.C.; Esteves,
F.A.; Lansac Toha, F.A.; Lima, A.F.; Influência do regime
hidrológico do rio Paraná sobre os valores da transparência,
oxigênio dissolvido e clorofila-a de três lagoas
de sua várzea. Revista Unimar, 14:153-162, 1992b (suplemento).
- Thomas, S.M.; Roberto, M.C.; Lansac
Toha, F.A.; Esteves, F.A.; Lima, A.F.; Dinâmica temporal
dos principais fatores limnológicos do rio Baía - planície
de inundação do Alto rio Paraná-MS, Brasil, Revista Unimar,
13(2):299-312, 1991.
- Thomas, S.M.; Roberto, M.C.; Lansac
Toha, F.A.; Lima, A.F.; Esteves, F.A. Características
limnológicas de uma estação de amostragem do Alto rio
Paraná e outras do baixo rio Ivinheima - (PR, MS-Brasil),
Acta Limnol. Brasil., 4:32-51, 1992c.
- Tundisi, J.G. Produção primária,
"standing-stock" e fracionamento do fitoplâncton na região
Lagunar de Cananeia. São Paulo, 1969, 131 p. (Tese, Universidade
de São Paulo)
|
|
|
