Voltar
INTRAPOPULATIONAL
VARIATION IN GROWTH RATES OF NESTLING RUDDY GROUND-DOVE
(Columbina talpacoti,Aves:Columbidae) IN BRAZIL
Renato Cintra1&
Roberto B. Cavalcanti2
1Departamento
de Ecologia do Instituto Nacional de Pesquisas da Amazônia-Manaus,AM-Brasil.
2Departamento
de Biologia Animal da Universidade de Brasília-Brasília,DF-Brasil.
ABSTRACT
Ricklefs (1968,1976)
suggested that tropical birds grow more slowly than birds
from temperate regions. However, this hypothesis has been
questioned (Oniki & Ricklefs 1981). The majority of
data on growth rates of neotropical birds are from Central
America and Surinam, with little data on intrapopulation
variation. In this paper,we focus intensively on one tropical
bird species (nestling Ruddy Ground-dove, Columbina talpacoti)
at one site in Brazil,and quantify the variation in growth
rates in relation to time of year, number of nestlings in
the nest, and hatching sequence. We used two methods to
estimate the growth parameters, that of Ricklefs(1967) and
that of Richards (Brisbin et al. 1986). Growth rate constants
for weight varied more among nests than within nests. Similar
variation was observed for asymptotic weight, and for final
wing length. Our results show that the Ruddy Ground-dove
is able to sustain nearly year-round reproductive activity,
with nestling growth rates much higher and more variable
than previously recorded for this species and tropical birds
in general. If variation within other tropical species proves
to be similar, it is clear that much larger sample sizes
will be required for meaningful interspecific comparisons.
Key words: nestling
growth rates; intrapopulational variation; birds, Ruddy
ground-dove, Columbina talpacoti.
INTRODUCTION
The Ruddy Ground-dove
(Columbina talpacoti) is widely distributed
in the Neotropical region. In Brasília, Central Brazil,
it breeds all year, affording the opportunity to study variation
in breeding parameters between the various seasons of the
year. In addition, several aspects of the breeding biology
of this common bird are yet unstudied (but see Haverschmidt
1953, Skutch 1956, Carvalho 1957, Trollope 1974).
The purpose of this
study is to examine the growth rates of nestling Ruddy Ground-dove.
Ricklefs (14, 15) suggested that tropical birds grow more
slowly than birds from temperate regions. However, this
hypothesis has been questioned (11). In Ricklefs' (15) study,
due to the scarcity of data, several species were represented
by data from only one nestling. The enormous variety of
tropical ecosystems may result in considerable variation
within the tropics, which could obscure comparisons between
tropical and temperate regions. The majority of data on
growth of neotropical birds are from Trinidad, Panama, and
Surinam, with little data on intrapopulation variation (11,
15). In this paper, we focus intensively on one tropical
species at one site in Brazil, and quantify the variation
in growth in relation to time of year, number of nestlings
in the nest and hatching sequence.
MATERIAL AND METHODS
The work was conducted
from January 1982 through February 1983 at the University
of Brasília (UnB) experimental farm, 20 km SE of Brasília
(15o 57 S and 47o 54 W), Brazil.
The study site comprised 6.5 ha, which included a variety
of native and introduced vegetation types. Native cerrado
vegetation (7, 12) covered 2.5 ha, a coffee plantation occupied
2 ha, and the remaining 2 ha included planted cypress (Cupressus
sp.) trees and a kitchen garden. The study was part of a
larger investigation on the reproductive biology of the
Ruddy Ground-dove (4, 5). To measure growth, we visited
nests daily between 0800 and 1000 am. In nests with two
nestlings, one was marked on the back with white acrylic
paint, to ensure individual identification. Body weight
as recorded to the nearest 0.2g using Pesola spring balances,
and wing length (curved wing) at 11 days (the time when
most fledglings were leaving the nest) as measured with
vernier calipers to within 0.1 mm. Nestlings were banded
with numbered aluminum bands(CEMAVE) on the right tarsus.
We used two methods to estimate the growth parameters, (1,
13). Use of the method proposed by Ricklefs(13) was undertaken
to compare our results with others in the literature. Therefore
most of the results and discussion sections are based on
parameters estimated using Ricklef's method. The Richard's
curve estimates an extra parameter (shape of the growth
curve) from the data. We used this method to test for shape
relationships which cannot be estimated by the Ricklef's
method and as a check for type II errors by the Ricklef's
logistic equations. Growth data for each nestling were fitted
to a logistic curve using the method of Ricklefs (13). The
equation had the form:
W(t) = A {1+exp[-K(t-ti)]}-1
where W(t) is the weight
(g) at age t (days), A is the asymptotic weight (g), K is
the growth rate constant (in days-1), and ti
is the age (days) at the point of inflection (11). We used
only weights that were less than 90% of the estimated asymptotic
weight.
Brisbin's procedures
comes from a reparameterized Richards process error sigmoid
growth model (1, 2). According to Brisbin et al. (1), the
procedure allows the independent quantification and statistical
comparison of three major characteristics of growth curves,"(1)size:
a measure of the magnitude of the asymptote approached by
the growth process; (2)rate: a measure of the approximate
amount of time required to complete growth; and (3)shape:
a value which expresses the specific path or trajectory
taken by the growth process to approach the asymptote within
the time constraints of the total growing period."
In this growth model
the equation has the following form:
(Wi+1-Wi
/ ti+1-ti) = (2(m+1) / T(1-m)) . (W¥1-mWim-Wi)+ei,
where Wi= body weight or size at time ti, W¥
= the asymptotic weight or size; T= the overall growing
time, indicative of growth rate; m= the Richards shape parameter;
and ei= the stochastic error at time ti.
Curve fitting procedures
to estimate the parameters W ,T and m which represents the
quantification of the characteristics
of growth size, rate
and curve shape respectively, were done using Non-linear
models from Systat (18). We also used Systat to run all
statistical analyses in this study.
RESULTS AND DISCUSSION
The Richard's model
was able to estimate an extra parameter, it did so at the
expense of precision in the estimates of asymptotic weight
(W) and overall growing time (T). The results from Tables
1 and 2 show that even though there were no significant
differences in the shape parameter, the Richards model was
not able to detect significant variation between treatments
in a asymptotic weight (W) shown by the "Ricklef's method".
Ruddy Ground-dove nestlings growth data (over 11 days period)
are consistent with a logistic curve (Fig. 1 A, B).
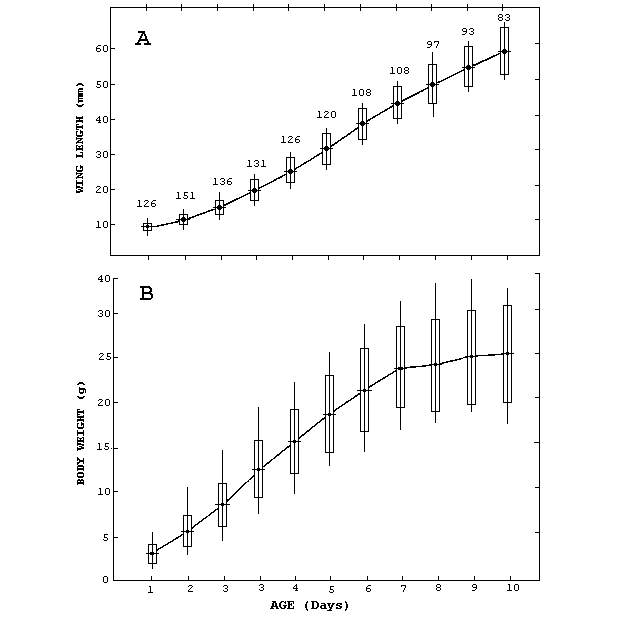
Figure 1.
Growth Curves for nestling Ruddy Ground-doves in relation
to age. (A) Wing length,(B) Body weight. Dots represent
means, rectangles are standard deviations, vertical lines
are ranges, and numbers are sample sizes.
One individual was banded
as a nestling and recaptured several times within the next
year. This bird left the nest with a weight of 30.5 g, 63%
of that of an adult male (Fig. 2B). Within three months
he had reached 97% of the mean adult male weight, and after
one year weighed 8% over the mean. He left the nest with
a wing length 71.4% of the male adults' mean (Fig. 2A).
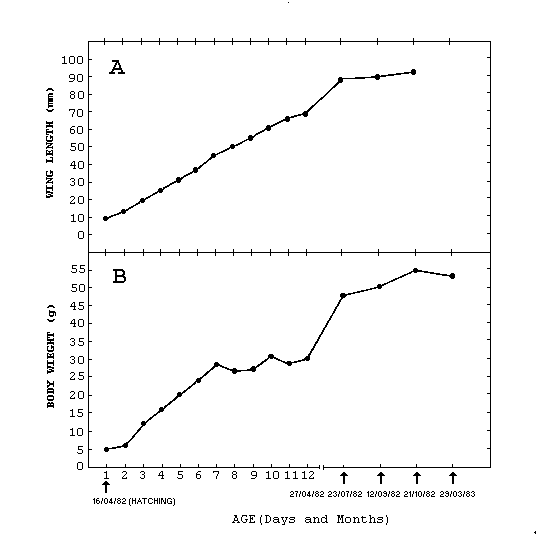
Figure 2.
Growth Curves in relation to age for one Ruddy Ground-dove
followed from nestling through its first year. (A) Wing
length,(B) Body weight.
The mean growth rate
constant for the population was 0.561 day-1 (SD= 0.113,
N= 111). Mean asymptotic weight was 27.8 g (SD= 3.105, N=111)
and mean final wing length was 59.2 mm (SD= 4.946, N= 107).
Growth rates were higher and asymptotic weights were similar
to those reported by Ricklefs (15) for this species. However,
the range of growth rate constants we observed (0.283 to
0.835) indicates substantial variation within the population,
and is comparable to the variation recorded among all species
surveyed by Ricklefs (15). Growth rate constants varied
more among nests (coefficient of variation= 16.6 %) than
within nests (CV= 8.32 %). Similar variation was observed
for asymptotic weight (CV among nests= 8.4 %, CV within
nests= 6.94 %), and for final wing length (CV among nests=
7.31 %, CV within nests= 3.99 %). Ruddy Ground-doves bred
throughout the year, adding a seasonal component to the
potential sources of variation in growth parameters. We
found significant variation in growth rate constants and
asymptotic weight between months (Tables 1 and 4), and variation
in final wing length approached significance (Tables 3 and
4). As expected, number of nestlings per nest had a significant
effect on both growth rate constants and asymptotic weight
(Table 1). Nests with one nestling produced birds with higher
growth rate constants and asymptotic weights. Final wing
length did not vary significantly between nests with one
and nests with two nestlings (Table 3). Thus, number of
nestlings per nest affected primarily rates of weight gain
rather than skeletal growth in the population studied. Interactions
between effects of month and number of nestlings per nest
were significant for growth rate constants, and approached
significance for asymptotic weight and final wing length
(Tables 1 and 3).
Table 1. Analysis
of variance of the growth rate constant, asymptotic weight
(using Ricklefs' method) in relation to month and number
of nestlings in the nest.
Ricklefs' growth rate constant (K)
| Source |
Sum
of Squares |
DF |
F |
Significance |
| Month |
0.196 |
4 |
4.801 |
0.001 |
| Nestlings |
0.075 |
1 |
7.395 |
0.008 |
| Interaction |
0.104 |
4 |
2.550 |
0.044 |
| ERROR |
1.029 |
101 |
|
|
Ricklefs asymptotic weight (A)
| Source |
Sum
of Squares |
DF |
F |
Significance |
| Month |
84.937 |
4 |
2.625 |
0.039 |
| Nestlings |
109.698 |
1 |
13.562 |
0.000 |
| Interaction |
61.607 |
4 |
1.904 |
0.116 |
| ERROR |
816.946 |
101 |
|
|
Table 2. Analysis
of variance of shape of growth curves, overall growing time
and asymptotic weight (Brisbin's proceedures), in relation
to month and number of nestlings in the nest.
Richard's shape of the growth curves
(m)
| Source |
Sum
of Squares |
DF |
F |
Significance |
| Month |
34.888 |
4 |
1.349 |
0.258 |
| Nestlings |
5.563 |
1 |
0.861 |
0.356 |
| Interaction |
10.055 |
4 |
0.389 |
0.816 |
| ERROR |
607.628 |
94 |
|
|
Richard's asymptotic weight
(W)
| Source |
Sum
of Squares |
DF |
F |
Significance |
| Month |
675.514 |
4 |
3.812 |
0.006 |
| Nestlings |
138.747 |
1 |
3.132 |
0.080 |
| Interaction |
146.538 |
4 |
0.827 |
0.511 |
| ERROR |
4163.981 |
94 |
|
|
Richard's overall growing time,indicative
of growth rate (T)
| Source |
Sum
of Squares |
DF |
F |
Significance |
| Month |
168.313 |
4 |
2.313 |
0.063 |
| Nestlings |
17.927 |
1 |
0.986 |
0.323 |
| Interaction |
34.334 |
4 |
0.472 |
0.756 |
| ERROR |
1709.768 |
94 |
|
|
Table 3. Analysis
of variance of the wing length at fledging, in relation
to month and number of nestlings in the nest.
| Source |
Sum
of Squares |
DF |
F |
Significance |
| Month |
193.030 |
4 |
2.190 |
0.076 |
| Nestlings |
20.062 |
1 |
0.911 |
0.342 |
| Interaction |
183.672 |
4 |
2.084 |
0.089 |
| ERROR |
2137.165 |
97 |
|
|
Table 4. Growth
rate constants (K),asymptotic weights(A),and wing length
at fledging(WING) for various times of year and nests with
one or two nestlings of Ruddy Ground-dove.
| Var |
#
of Nestl. |
Feb |
Abr |
May |
Jul |
Nov |
Total |
| K |
One |
0.451 |
0.680 |
0.616 |
0.669 |
0.706 |
0.617
a |
|
|
(0.106) |
(0.010) |
(0.099) |
(0.122) |
(0.064) |
(0.129)
b |
|
|
5 |
4 |
4 |
6 |
3 |
22
c |
|
|
|
|
|
|
|
|
|
Two |
0.512 |
0.537 |
0.569 |
0.515 |
0.637 |
0.547 |
|
|
(0.139) |
(0.077) |
(0.112) |
(0.l01) |
(0.076) |
(0.104) |
|
|
8 |
24 |
32 |
20 |
5 |
89 |
|
|
|
|
|
|
|
|
| A |
One |
32.4 |
31.5 |
30.9 |
28.1 |
26.2 |
29.9 |
|
|
(2.82) |
(3.03) |
(1.41) |
(3.35) |
(3.25) |
(3.44) |
|
|
5 |
4 |
4 |
6 |
3 |
22 |
|
|
|
|
|
|
|
|
|
Two |
27.4 |
27.0 |
27.5 |
27.1 |
26.7 |
27.2 |
|
|
(5.49) |
(2.87) |
(2.01) |
(2.55) |
(2.49) |
(2.78) |
|
|
8 |
24 |
32 |
20 |
5 |
89 |
|
|
|
|
|
|
|
|
| WING |
One |
57.9 |
63.0 |
59.8 |
57.0 |
59.3 |
59.2 |
|
|
(5.78) |
(2.83) |
(1.93) |
(7.66) |
(3.18) |
(5.16) |
|
|
5 |
4 |
4 |
5 |
2 |
20 |
|
|
|
|
|
|
|
|
|
Two |
53.8 |
59.1 |
59.5 |
61.5 |
56.9 |
59.2 |
|
|
(7.49) |
(4.68) |
(3.48) |
(3.36) |
(9.07) |
(4.93) |
|
|
8 |
24 |
32 |
19 |
4 |
87 |
a= mean; b=standard
error; c= sample size.
The first-hatched nestlings
had significantly higher final wing lengths than second-hatched
nestlings (Mann-Whitney U-Test, U = 1003, p<0.05, N=80).
Growth rate constants (K) and asymptotic weights (A) were
not significantly different between first- and second-hatched
nestlings (K: Mann-Whitney U-Test, U=794, p=0.95, N=80;
A: Mann-Whitney U-Test,U=797, p=0.97, N=80). Growth rate
constants (K) and asymptotic weight (A) were not significantly
correlated (r2= -0.07, p>0.05, N=111). However,
final wing length showed low but significant correlation
with both K (r2=-0.223, p<0.05, N=107) and
A (r2=0.189, p<0.05, N=107).
Growth rates in the
Ruddy Ground-dove in Brasília are much higher than reported
in previous studies. Ricklefs (15), based on a sample of
four nestlings from Surinam (8), found a mean of K= 0.460
days-1. In our sample, 77% of the birds had K's above 0.460
days-1, and 55% had K's above the overall mean of 0.520
days-1 estimated for neotropical birds (Ricklefs 15). This
result may be in part a consequence of the situation studied.
The granivorous ground-doves had a large supply of seeds
and farm grains(6), and nest predation was high (5). As
suggested by Ricklefs (14), high growth rates may be more
advantageous where there
is strong pressure to leave the nest early, where parents
produce several clutches in a year, or where there is high
predation in the nest. High growth rates can only be realized
however in situations
of ample food supply, to meet the demands of the nestlings.
This combination of factors may well be occuring in the
population we studied. Our data show that there is considerable
plasticity in growth rates within this species, allowing
for quick environmental responses. For example, growth rates
and asymptotic weights were higher in nests with one nestling
than in nests with two nestlings. Similar declines in growth
parameters with increasing brood size have been reported
for other columbids, such as the Woodpigeon,Columba palumbus,
and the Mourning Dove,Zenaida macroura (9, 10).
Our data are in agreement
with evidence that variation in growth parameters is smaller
within nests than between nests, possibly because nestlings
in the same nest are exposed to more uniform conditions
than in population as a whole, and/or have more similar
genotypes (15).
Our results show that
the Ruddy Ground-dove is able to sustain nearly year-round
reproductive activity, with nestling growth rates much higher
and more variable than previously recorded for this species
and tropical birds in general. If variation within other
tropical species proves to be similar, it is clear that
much larger sample sizes will be required for meaningful
interspecific comparisons. Most previous growth data on
neotropical birds comes from Central America. Further studies
of South American species, especially those of open habitats,
may yield a more general model of patterns of variation
in growth characteristics in tropical landbirds.
ACKNOWLEDGMENTS
This research benefitted
from the help of many people. Field assistance was provided
by Regina Macedo, Maria Alice. Alves, Mauro Valle, Adriana
Moreira, Rodrigo Cavalcanti, and the workers at the University's
Experimental Farm. Dr. Colin E. Johnson provided banding
equipment and advice. Bird bands used were from CEMAVE,
through Paulo Antas. An earlier version of the manuscript
was criticized by Edwin Willis and Thomas E. Lacher. Alvaro
Negret, Raimundo Henriques and William Magnusson also gave
useful suggestions. Revision by Tânia Sanaiotti was very
much appreciated. The Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) supported the work.
RESUMO
Variação intrapopulacional
nas taxas de crescimento de filhotes de Rolinha Caldo-de-Feijão
(Columbina talpacoti, Aves:Columbidae) no Brasil.
Ricklefs (1968,1976) sugeriu que aves tropicais crescem
mais lentamente do que aves de regiões temperadas. Contudo,
essa hipótese tem sido questionada (Oniki & Ricklefs
1981). A maioria dos dados sobre taxa de crescimento de
aves neotropicais são da América Central e Suriname, com
poucos dados sobre variação intrapopulacional. Neste artigo,
nós concentramos intensivamente nossas observações em uma
espécie tropical (filhotes
de Rolinha Caldo-de-Feijão, Columbina talpacoti)
para um local no Brasil, e quantificamos a variação nas
taxas de crescimento em relação ao período do ano, número
de filhotes no ninho, e a sequência de eclosão.
Nós usamos dois métodos
para estimar os parâmetros de crescimento, o de Ricklefs(1967)
e o de Richards (Brisbin et al.1986). As constantes da taxa
de crescimento para peso variaram mais entre ninhos que
dentro dos ninhos. Variação similar foi observada para o
peso assintótico, e para o comprimento final da asa. Nossos
resultados mostram que a Rolinha Caldo-de-Feijão é capaz
de manter atividade reprodutiva por quase o ano inteiro,
com taxas de crescimento de filhotes muito maiores e mais
variáveis que o registrado antes para esta espécie e para
aves tropicais em geral. Se a variação dentro de espécies
tropicais for similar, está claro que tamanho de amostras
muito maiores serão necessárias para comparações interespecíficas
significativas.
REFERENCES
- Brisbin,I.L.Jr.;White,G.C.;Bush,P.B.1986."PCB
intake and the growth of waterfowl:multivariate analyses
based on a reparameterized Richards sigmoid model"Growth
50:1-11.
- Brisbin,I.L.Jr.; K.W.McLeod,K.W.;
G.C.White. 1987. Sigmoid growth and the assessment of
Hormesis:a case for caution. Health Physics 52(5):553-559.
- Carvalho,C.1957. Relações biológicas
entre Columbigallina passerina e C.talpacoti
(Aves, Columbidae).Bol.Mus.Par. Emílio Goeldi 7:1-15.
- Cintra,R.1983.Taxas de crescimento
de filhotes de rolinha, Columbina talpacoti Aves:
Columbidae, em relação a fatores ecológicos e populacionais
no Planalto Central. Unpubl. M. S. Dissertation, Universidade
de Brasília,Brasília,Brasil.114 pp.
- Cintra,R.1988. Reproductive ecology
of the Ruddy Ground-dove, Columbina talpacoti (Columbidae),in
the Central Plateau of Brazil.Wilson Bull.100(3):443-457.
- Cintra,R.; Alves,M.A.S.; Cavalcanti,R.B.
1990. Dieta da Rolinha Columbina talpacoti(Aves,
Columbidae) no Brasil Central-Comparação entre sexos e
idades.Rev.Brasil. Biologia.50(2):469-473.
- Eiten,G. 1972. The cerrado vegetation
of Brasil.Bot.Rev. 38:201-341.
- Haverschmidt,F.1953.Notes on the
life history of Columbigallina talpacoti in Surinam.Condor
55: 21-25.
- Holcomb, L. C.; Jaeger,M. 1978. Growth
and calculation of age in Mourning Dove nestlings.J.Wild.Manage.42:843-852.
- Murton,R.K.; Isaacson,A.J.; Westwood,N.J.
1963. The food and growth of nestling woodpigeons in relation
to the breeding season. Proc. Zool. Soc. London 141:747-781.
- Oniki,Y.; Ricklefs,R.E.1981. More
growth rates of birds in the humid New World tropics.Ibis
123: 349-354.
- Ratter,J. 1980.Notes on the vegetation
of Fazenda Água Limpa, Brasília, DF.Royal Botanic Garden,
Edinburgh, Additional series,No.1,124p.
- Ricklefs, R.E.1967. A graphical method
of fitting equations to growth curves. Ecology 48: 978-983.
- Ricklefs,R.E.1968. Patterns of growth
in birds. Ibis 110:419-451.
- Ricklefs,R.E.1976.Growth rates of
birds in the humid New World tropics.Ibis 118:179-207.
- Skutch,A.F.1956. Life history of
the Ruddy Ground-dove. Condor 58:188-205.
- Trollope,J.1974. The breeding and
behaviour of the Talpacoti dove. Avicult. Mag.80:86-92.
- Wilkinson,L.1991. SYSTAT: The System
for Statistics. SYSTAT, Inc. Evanston.
