Voltar
EFEITO DA MINERAÇÃO DE AREIA SOBRE A
ESTRUTURA DA COMUNIDADE DE MACRÓFITAS AQUÁTICAS EM UM ECOSSISTEMA
LÓTICO DO LITORAL SUL PAULISTA.
Antonio Fernando Monteiro
Camargo1 Alexandre Schiavetti2,3 &
Maurício Cetra3
1Departamento
de Ecologia/Caunesp, Instituto de Biociências, Unesp, C.P.
199, 13506-900, Rio Claro, SP, Brasil.
2 Bolsista
DTI - CDCC - USP - São Carlos.
3Programa
de Pós-graduação em Ciências da Engenharia Ambiental - CHREA
- USP - São Carlos.
RESUMO
Neste trabalho nós avaliamos
a estrutura da comunidade de macrófitas aquáticas em dois
rios da bacia do Rio Itanhaém, Rio Branco e Rio Preto (litoral
Sul Paulista) em três épocas do ano. Foram realizadas coletas
de macrófitas aquáticas em oito porções do leito dos dois
rios, localizados na planície costeira, e medida a biomassa
de cada espécie. A partir dos valores de biomassa foram
calculados os índices de diversidade de Shannon e Simpson,
os índices de uniformidade, dominância e similaridade. Foram
observadas diferenças na ocorrência das espécies e nos valores
de biomassa entre os dois rios e as épocas de coleta. Os
índices de estrutura de comunidades mostraram uma forte
dominância de Eichhornia azurea no R. Branco, enquanto
no R. Preto equitabilidade na biomassa. A diversidade de
espécies é maior no R. Preto, devido a homogeneidade de
distribuição de biomassa entre as espécies e o maior número
de espécies. As diferenças observadas na estrutura da comunidade
de macrófitas aquáticas nos dois rios, provavelmente, se
deve a intensa atividade de mineração de areia no R. Branco,
que provocou mudanças no leito deste rio em um intervalo
de aproximadamente 25 anos.
Palavras chave:
macrófita aquática, estrutura de comunidade, mineração de
areia, bacia do Rio Itanhaém.
INTRODUÇÃO
Macrófitas aquáticas
desenvolvem-se abundantemente em ecossistemas rasos, de
baixa turbulência e com amplas áreas marginais, ocorrendo
em estandes mistos ou monoespecíficos (3). Esta vegetação
desempenha um importante papel nestes ecossistemas, influenciando
aspectos físicos e químicos da água, servindo como substrato
para o perifíton, como local de abrigo e fonte de alimento
para invertebrados aquáticos e como importantes produtores
primários do ecossistema (5).
Em ambientes lênticos,
submetidos ao processo de eutrofização artificial, macrófitas
flutuantes proliferam abundantemente, prejudicando a navegação,
o lazer, a utilização da água para geração de energia elétrica
e para abastecimento (6). A construção de represas e açudes
também pode promover o desenvolvimento de macrófitas aquáticas.
O barramento de rios e córregos pode resultar na formação
de ambientes aquáticos com baixa turbulência, amplas regiões
marginais e com grande transparência, propiciando o desenvolvimento
de macrófitas aquáticas emersas, com folhas flutuantes e
submersas (10). Estas últimas podem causar grandes problemas
à navegação, prejudicar a pesca e a geração de energia elétrica.
A mineração de areia
em leito de rios, modifica a morfologia destes, a médio
e longo prazo, pela retirada de areia e o desbarrancamento
das margens, podendo criar ambientes favoráveis ao crescimento
de macrófitas. Desta forma, a estrutura da comunidade de
macrófitas em um rio submetido à atividade de mineração
de areia pode ser alterada. Alterações na estrutura de comunidades
podem ser evidenciadas através do índice de diversidade
de espécies, que segundo Magurran (7), freqüentemente é
visto como um bom indicador da integridade dos sistemas
ecológicos.
Neste trabalho nós obtivemos
dados sobre o índice de diversidade de espécies da comunidade
de macrófitas em dois rios da bacia do Rio Itanhaém (litoral
sul Paulista - Brasil), um deles submetido a intensa mineração
de areia e outro não submetido à esta atividade. O objetivo
principal deste trabalho é verificar como a atividade de
mineração de areia pode alterar a estrutura da comunidade
de macrófitas aquáticas, indicando o grau de integridade
do ecossistema.
ÁREA DE ESTUDO
O litoral centro-sul
Paulista vem sofrendo uma intensa ocupação humana a cerca
de 30 anos, em função de sua vocação turística e de sua
proximidade e facilidade de acesso à partir da capital do
Estado. Residências de veraneio ocupam praticamente toda
a orla marítima e desenvolvem-se atualmente para o interior,
formando um grande conglomerado urbano unindo os municípios
de Praia Grande, Mongaguá, Itanhaém e Peruíbe. Paralelamente
à rápida expansão urbana, intensifica-se a atividade de
mineração de areia em rios da região.
No município de Itanhaém
localiza-se a maior parte da bacia hidrográfica do Rio Itanhaém
(23o 50', 24o 15' S e 46o
35', 47o 00' W) que possui aproximadamente 50
% de sua área localizada na planície costeira (Figura 1).
A planície apresenta baixa declividade, com altitudes de
apenas 3,0m a 6,0km da costa e nesta parte da bacia os rios
apresentam leitos meândricos, áreas alagadas, braços abandonados
e baixa velocidade de corrente. Estas características propiciam
a ocorrência abundante de macrófitas aquáticas de diferentes
espécies e tipos ecológicos, tanto nos canais dos rios,
como em áreas alagadas.
Os rios Preto (águas
pretas) e Branco (águas brancas) são formadores do Rio Itanhaém,
percorrem grandes extensões da planície costeira e apresentam,
em áreas de remanso, grande quantidade de macrófitas aquáticas
flutuantes, emersas, submersas e com folhas flutuantes.
No Rio Branco a atividade de mineração de areia é intensa
em um trecho de aproximadamente 3km, enquanto no Rio Preto
esta atividade não é realizada.
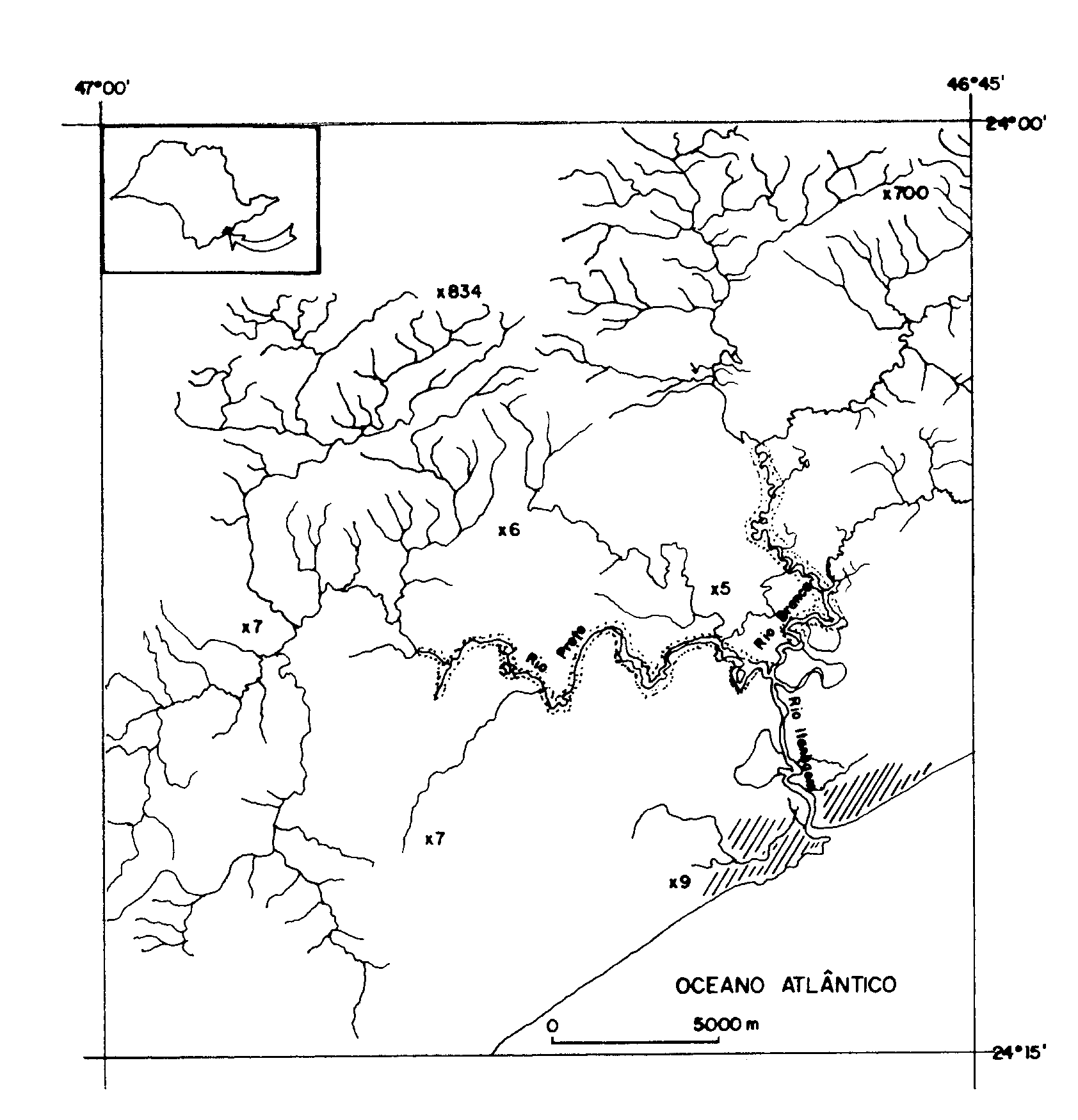
Figura 1.
Localização da área de estudo no Estado de São Paulo e na
Bacia do Rio Itanhaém (áreas pontilhadas).
METODOLOGIA
Coletas de macrófitas
aquáticas foram realizadas nos rios Preto e Branco em três
períodos do ano (maio, julho e setembro/91), na região de
ambos os rios localizada na planície costeira. Os trechos
amostrados foram divididos em 8 porções, nas quais obteve-se,
em cada uma, amostras de macrófitas (em duplicata) através
de um quadrado vazado de 0,25m2. Após a coleta,
as diferentes espécies foram separadas e posteriormente
secas em estufa (80°C) até peso constante. A biomassa de
cada espécie, em cada rio, foi calculada a partir da média
das replicas e transformada para grama de peso seco / m2
. Os índices de diversidade de Shannon Simpson (8) foram
calculados através da biomassa das espécies de macrófitas
aquáticas existentes nos trechos estudados dos rios Preto
e Branco, utilizando-se as seguintes fórmulas:
Índice de Shannon
_
H = - å
( ni/N) Log (ni /N)
e Índice de Simpson
S = 1 - C e
C = å
ni (ni - 1)/ N (N - 1)
onde ni =
valor de biomassa para cada espécie
N= total de biomassa
Aos valores dos índices
de diversidade obtidos foi aplicada a técnica "Jack-knife"(7),
para estimar os intervalos de confiança dos índices de diversidade.
Os índices de uniformidade
(9) , dominância (11) e de similaridade (12) também foram
calculados baseados nos dados de biomassa
_
Uniformidade x = H/ Log
S
onde S = nº de espécies
Dominância C = å (ni/N)2
Similaridade S = 2C/a
+ b
onde a = nº de espécies
encontrada na comunidade A;
b = nº de espécies encontrada
na comunidade B e;
c = nº de espécies comuns
às duas comunidades.
As modificações na morfologia
do leito do Rio Branco, ocasionadas pela atividade de mineração
de areia foram obtidas à partir da superposição de duas
cartas geográficas da região, elaboradas à partir de fotos
aéreas com um intervalo de 24 anos. A carta Itanhaém, IBGE,
1971 (fotos aéreas de 1962), foi superposta a carta Itanhaém,
IGC, 1988 (fotos aéreas de 1986/1987). À partir da superposição
das cartas foi elaborado o mapa que mostra as alterações
provocadas no leito do R. Branco pela atividade de mineração
de areia.
RESULTADOS
A Tabela 1 apresenta
a lista de espécies de macrófitas aquáticas e os valores
de biomassa de cada espécie em cada época do ano que foram
amostradas nas áreas estudadas dos rios Preto e Branco.
Observa-se pela tabela que cinco espécies são comuns aos
dois rios, duas espécies exclusivas do Rio Preto e uma espécie
exclusiva do Rio Branco, considerando-se as três épocas
de coleta.
Tabela 1. Biomassa
das espécies de macrófitas aquáticas amostradas nos rios
Branco e Preto nos meses de maio, julho e setembro.
| |
maio |
julho |
Setembro |
| Rio
Branco |
|
|
|
| Eichhornia
azurea |
528,32 |
297,82 |
336,53 |
| Eichhornia
crassipes |
298,82 |
46,98 |
0,00 |
| Anacharis
densa |
12,39 |
47,48 |
55,77 |
| Salvinia
molesta |
11,21 |
17,19 |
4,71 |
| Pistia
stratiotes |
29,21 |
1,63 |
3,30 |
| Utricularia
foliosa |
0,00 |
0,00 |
0,04 |
| Gramineae |
2,20 |
0.0 |
1,92 |
|
|
|
|
| Rio
Preto |
|
|
|
| Eichhornia
azurea |
75,41 |
67,75 |
171,32 |
| Eichhornia
crassipes |
290,95 |
63,31 |
58,70 |
| Anacharis
densa |
5,25 |
29,45 |
76,39 |
| Ricciacarpus
natans |
0,00 |
0,00 |
2,84 |
| Salvinia
molesta |
38,39 |
12,51 |
50,59 |
| Utricularia
foliosa |
0,94 |
0,83 |
0,74 |
| Nymphaea
rudgeana |
5,00 |
12,51 |
0,00 |
| Polygonum
sp |
31,98 |
0,00 |
0,00 |
| Cyperus
sp |
10,22 |
26,74 |
105,31 |
| Gramineae |
0,84 |
55,97 |
0,00 |
Observando-se os resultados
ficam evidentes as diferenças de biomassa entre espécies
de um mesmo rio, como por exemplo no Rio Branco em maio
(528,32g PS.m-2 para E. azurea e 11,21g
PS. m-2 para S. molesta). Diferenças entre
épocas do ano também podem ser observadas, tal como, E.
crassipes no Rio Preto (290,95g PS.m-2 em
maio e 58,70g PS.m-2 em setembro). Comparando
os
dois rios estudados
observa-se que no Rio Branco, em todas as épocas do ano,
E. azurea apresenta valor de biomassa sempre muito
superior às outras espécies, enquanto no Rio Preto existe
um maior equilíbrio na distribuição de biomassas entre as
espécies. Os índices de dominância, uniformidade e de similaridade
evidenciam melhor as diferenças entre as comunidades de
macrófitas aquáticas dos dois rios. Os índices de uniformidade
em maio (0,32), julho (0,32) e setembro (0,28) no Rio Branco,
foram inferiores aos calculados em maio (0,54), julho (0,85)
e setembro (0,74) no Rio Preto, indicando que neste último
há uma maior uniformidade na comunidade de macrófitas. Os
índices de dominância também evidenciam as diferenças entre
a comunidade de macrófitas aquáticas dos dois
rios. No Rio Branco
os índices de dominância foram de 0,74 em maio, 0,54 em
julho e 0,72 em setembro, enquanto no R. Preto de 0,43 em
maio e 0,25 em julho e setembro. Os índices de similaridade
calculados para as três épocas do ano mostram que as comunidades
dos dois rios são similares (0,71 em maio e julho e 0,53
em setembro). Os índices calculados mostram também que as
comunidades dos dois rios apresentam uma grande dinâmica
temporal pois apresentam valores maiores ou menores dependendo
da época do ano. Na tabela 2 são apresentados os índices
de diversidade de Shannon e Simpson, os valores do "Jack-knife"
e os intervalos de confiança.
Tabela 2. Valores
dos índices de diversidade de Shannon (H) e Simpson (Ds),
"Jack-knifes" (JK) e intervalos de confiança (IC) das comunidades
de macrófitas aquáticas dos rios Branco e Preto em diferentes
meses do ano. *índice de diversidade dentro do intervalo
de confiança.
| |
|
Rio
Branco |
|
|
Rio
Preto |
|
|
|
|
|
|
|
|
| meses |
H |
JK |
IC |
H |
JK |
IC |
|
|
|
|
|
|
|
| maio |
0,592 |
0,615 |
0,594-0,637 |
1,188* |
1,199 |
1,163-1,124 |
| julho |
0,947 |
1,000 |
0,955-0,1052 |
1,792 |
1,995 |
1,925-2,064 |
| setembro |
0,545* |
0,584 |
0,518-0,649 |
1,534* |
1,740 |
1,714-1,766 |
|
|
|
|
|
|
|
|
Ds |
JK |
IC |
Ds |
JK |
IC |
|
|
|
|
|
|
|
| maio |
0,252* |
0,253 |
0,249-0,258 |
0,558* |
0,553 |
0,546-0,561 |
| julho |
0,464* |
0,474 |
0,456-0,492 |
0,811 |
0,956 |
0,952-0,845 |
| setembro |
0,281 |
0,233 |
0,197-0,270 |
0,776 |
0,843 |
0,840-0,845 |
Os dois índices de diversidade
calculados mostram que o Rio Preto apresenta maior diversidade
de espécies de macrófitas aquáticas quando comparado com
o rio Branco. Os valores calculados para a diversidade de
espécies estão sempre próximos aos obtidos ou dentro do
intervalo de confiança encontrado pela técnica "Jack-knife",
demonstrando a possibilidade de se utilizar este índice
na comparação da estrutura da comunidade de macrófitas nestes
ambientes.
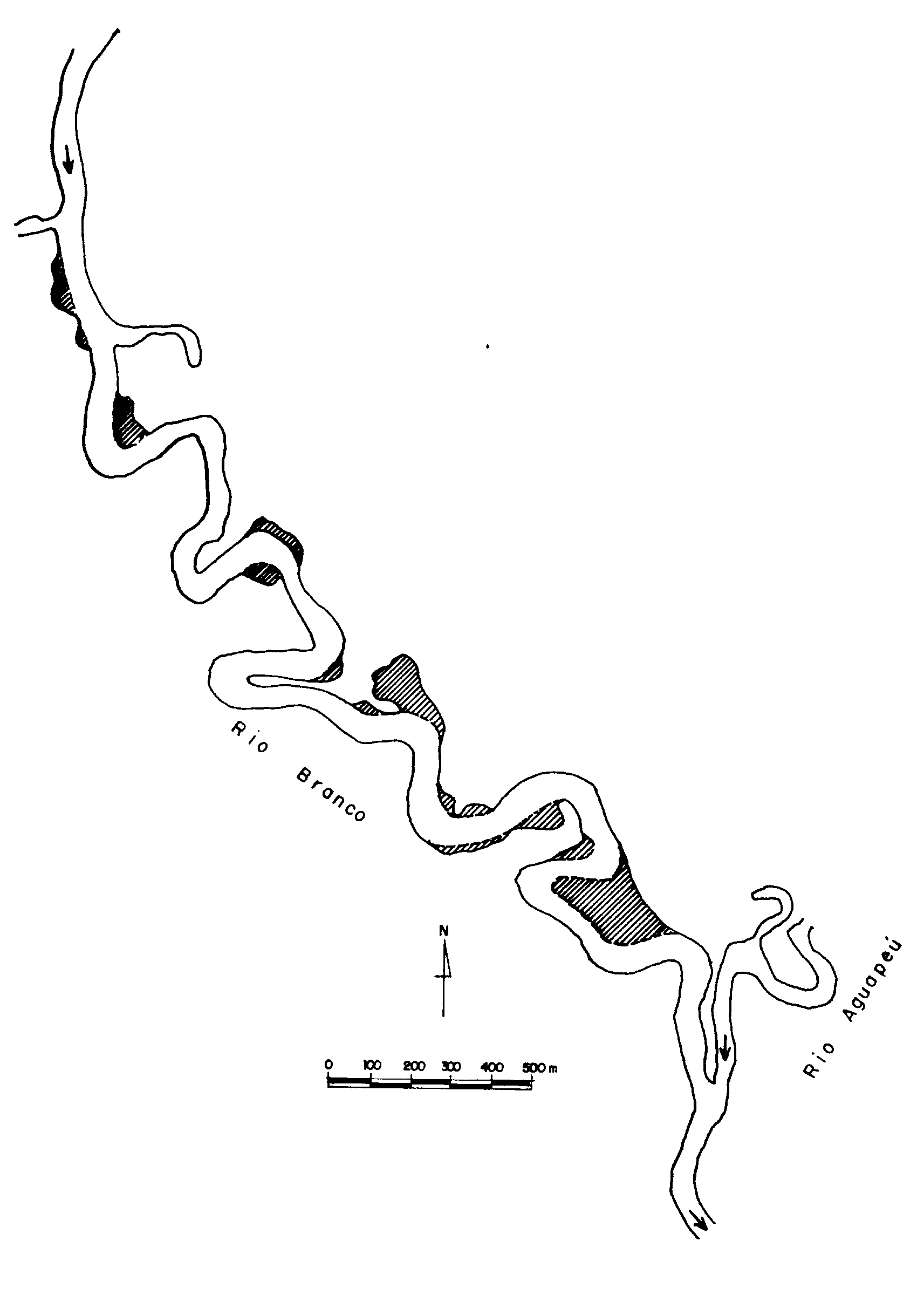
A Figura 2 mostra as
áreas criadas antropicamente no leito do Rio Branco pela
atividade de mineração de areia. As áreas em branco referem-se
à morfologia do leito do rio no ano de 1962, ou seja o leito
original do Rio Branco, enquanto em hachureado estão destacadas
as áreas formadas pela atividade de mineração de areia até
o ano de 1986/87, que corresponde aproximadamente à morfologia
atual do leito do rio. O leito do Rio Preto não sofreu nenhuma
modificação durante este mesmo período de tempo.
Figura 2. Trecho
do rio Branco mostrando as áreas criadas antropicamente
pela atividade de mineração de areia (partes hachureadas).
DISCUSSÃO
As comunidades
de macrófitas aquáticas, embora freqüentemente sejam fisionomicamente
similares, apresentam grandes diferenças na sua estrutura
devido a diversos fatores, tais como, natureza físico-química
do substrato, profundidade da coluna dágua, disponibilidades
de nutrientes e as interações entre as diferentes populações
(13). As comunidades dos Rios Preto e Branco são fisionomicamente
semelhantes em todas as épocas do ano, fato que se deve
à presença abundante de E. azurea, S. molesta
e A. densa nos dois rios. Poucas espécies, e com
biomassa reduzida, são exclusivas do R. Preto, tais como,
N. rudgeana, Ricciacarpus sp, Polygonum
sp e Cyperus sp. Apesar desta aparente semelhança
entre as duas comunidades, os resultados obtidos demonstram
a grande diferença na estrutura e dinâmica de biomassa das
comunidades de macrófitas aquáticas, pois no R. Branco observa-se
forte dominância de E. azurea, enquanto isto não
ocorre no R. Preto. Esta dominância é corroborada pela análise
dos valores de uniformidade calculados, pelos quais observa-se
grande concentração de biomassa em uma única espécie no
R. Branco (E. azurea) e uma equitabilidade de biomassa
entre as espécies do R. Preto. As diferenças na estrutura
das comunidades de macrófitas aquáticas dos dois rios estudados
poderiam ser produto das diferenças de características físico-químicas
da água, no entanto, os rios Preto e Branco embora apresentem
aspectos de água distintos,
possuem características físico-químicas similares (1, 4).
A criação de áreas de remanso no Rio Branco, pela atividade
de mineração de areia, evidenciadas pela superposição das
cartas elaboradas à partir de fotografias aéreas de 1962
e 1986/87 sugerem que as diferenças observadas na estrutura
da comunidade de macrófitas aquáticas dos dois rios se devem,
provavelmente, às modificações morfológicas do leito do
Rio Branco. De fato, Magurran (7) destaca que ambientes
submetidos a fortes impactos antrópicos apresentam mudanças
nas relações espécie-abundância de suas comunidades. A menor
diversidade de espécies e a dominância de E. azurea
no R. Branco, provavelmente, estão associadas à influência
antrópica neste rio (mineração de areia) que promoveu o
surgimento de locais propícios à instalação de espécies
de macrófitas
aquáticas com largo
espectro ecológico, tal como, E azurea. Camargo &
Esteves (2) evidenciaram o largo espectro ecológico desta
espécie em uma lagoa marginal do Rio Mogi-Guaçu que é submetida
a uma ampla variação de nível dágua.
A atividade de mineração
de areia por um período de mais de vinte anos, no R. Branco,
criou áreas de pouca correnteza e de baixa profundidade
(áreas de remanso), propícias à colonização de macrófitas
aquáticas. E. azurea, uma espécie abundante em rios
da bacia do R. Itanhaém, e de largo espectro ecológico,
colonizou estas áreas, alterando a estrutura da comunidade
de macrófitas aquáticas e evidenciando o efeito da ação
antrópica no R. Branco.
AGRADECIMENTOS
Os autores agradecem
o CAUNESP (Centro de Aquicultura da UNESP) e o Depto de
Ecologia UNESP/Rio Claro pelas facilidades e ao CNPq pela
Bolsa de Iniciação Científica. Aos Mestres Luís Maurício
Bini e Silmara de Cássio Luciano, ao Técnico Carlos Fernando
Sanches pelas ajudas em campo e em laboratório e ao Prof.
Dr. Harold Fowler pela tradução do abstract.
ABSTRACT
Effect of sand extration
on aquatic macrophyte community structure in a lotic ecosystem
of the coast region of São Paulo State. In this paper
we evaluate aquatic macrophyte community structure in two
rivers, Rio Branco and Rio Preto, in the Itanhaém River
basin of the coastal region of São Paulo, in three different
periods of the year. Aquatic macrophytes were collected
in eight portions of the river bed in the two rivers, located
in the coastal plain, and the biomass of each species was
measured. From biomass values, the indices of Shannon e
Simpson, uniformity, dominance and similarity were calculated.
Differences in species composition and these indices were
found between
rivers and collection
dates. Community struture indices demonstrated a strong
dominance of Eichhornia azurea in the Rio Branco,
while the Rio Preto was characterized by biomass equitability.
Species diversity was greater in the Rio Preto, due to the
heterogeneity of species biomass distributions and a greater
number of species. The observed differences in the aquatic
macrophyte community strutures in these two rivers is probably
due to sand extration industry in the Rio Branco, which
caused changes in the river bed over a 25 year period.
Key words: aquatic
macrophytes, community struture, sand extration, Itanhaém
River basin.
REFERÊNCIAS
- Camargo, A.F.M.; Bini, L.M.; Schiavetti,
A. & Ferreira, R.A.R. Influencia de la fisiografia
y de las actividades humanas en las características limnológicas
de rios del litoral Sur paulista - Brasil. Tankay,
1: 275 276,1994.
- Camargo, A.F.M.; Esteves, F.A. Influence
of water level variation on biomass and chemical composition
of the aquatic macrophyte Eichhornia azurea (Kunt)
in an oxbow lake of the Rio Mogi-Guaçu (São Paulo, Brasil).
Arch. Hydrobiol. 135(3): 423-432, 1996.
- Camargo, A.F.M.; Esteves, F.A.. Influence
of water level variation on fertilization of an oxbow
lake of Rio Mogi-Guaçu, state of São Paulo, Brasil. Hydrobiologia,
299: 185-193, 1995.
- Camargo, A.F.M.; Ferreira, R.A.R.;
Schiavetti, A.; Bini, L.M. Influence of physiography and
human activity on limnological characteristics of lotic
ecosystems of the south coast of São Paulo, Brazil. Acta
Limnol. Brasil.:231-243, 1996.
- Esteves, F. A. Fundamentos de
Limnologia. Interciência/FINEP, Rio de Janeiro,
1986.
- Gopal, B. Ecology and Management
of Aquatic Vegetation in the Indian Subcontinent.
Kluwer Academic Publishers, Dordrecht, 1990.
- Magurran, A.E. Ecological diversity
and its measurement. University Press, Cambridge,
1988.
- Odum, E.P. Fundamentals of Ecology.
W.B. Saunders Company, Philadelphia, 1971.
- Pielou, E.C. The measurement of diversity
in different types of biological colletions. J. Theoret.
Biol., 13: 131-144, 1966.
- Pieterse, A. H.; Murphy, K.J. Aquatic
Weeds: The Ecology and Management of Nuisance Aquatic
Vegetation. Oxford University Press, New York, 1990.
- Simpson, E.H. Measurement of diversity.
Nature, 163: 688, 1949.
- Sorensen, T. A method of establishing
groups of equal amplitude in plant society based on similarity
of species content. K. Danske Vidensk Selsk.
5: 1-34, 1948.
- Zutshi, D. P.; Gopal, B. Struture
and dynamics of plant communities. In: B. Gopal (ed.),
Ecology and Management of Aquatic Vegetation in the
Indian Subcontinent. Kluwer Academic Publishers, Dordrecht,
127-146, 1990.
