|
|
ARTIGOS I Fórum de Debates ECOLOGIA DA PAISAGEM E PLANEJAMENTO AMBIENTAL |
|
|
|
ARTIGOS I Fórum de Debates ECOLOGIA DA PAISAGEM E PLANEJAMENTO AMBIENTAL |
|
|
DELIMITAÇÃO PRELIMINAR DOS BIÓTOPOS NATURAIS NA BACIA DO ARROIO DILÚVIO, VISANDO CONTRIBUIR PARA O MANEJO DOS RECURSOS HÍDRICOS EM PORTO ALEGRE, RS.
Maria Luiza Porto1; Heinrich Hasenack1; Paulo Brack2; Sérgio Luis Carvalho Leite2; Eduardo Dias Forneck3; Rafael Garziera Perin3; Taís Cristine Ernst Frizzo3 1 Departamento de Ecologia, Instituto de Biociências, UFRGS2 Departamento de Botânica, Instituto de Biociências, UFRGS3 Programa de Pós-Graduação em Ecologia, UFRGS
INTRODUÇÃO A região da grande Porto Alegre-RS vem sofrendo intenso risco de destruição de sua biodiversidade devido ao acelerado avanço urbanístico, seja este irregular ou regular. Os ecossistemas originais vão dando lugar a loteamentos, muitas vezes clandestinos, não existindo políticas públicas de proteção à natureza ou, quando existem, não são eficientes. As áreas naturais exercem importância significativa sobre a qualidade ambiental do município através de seu valor paisagístico, biodiversidade, estabilidade climática, hidrologia e controle de erosão, entre outros aspectos. A capital do estado, Porto Alegre, possui ainda cerca de 4.500ha de matas nativas em seus morros (Porto e Oliveira, 1998). Este município apresentou, em seu território, cerca de 170 espécies arbóreas nativas (Brack et al., 1998), o que representa, aproximadamente, um terço das espécies que ocorrem no estado. Na execução deste trabalho, estão sendo utilizados recursos avançados de geoprocessamento. Imagens de satélite e métodos atuais estão servindo como ferramentas para identificação de atributos da paisagem, de forma a levar a uma visão global e sintética dos problemas ambientais e orientar soluções adequadas ao manejo e gerenciamento de áreas no âmbito do município de Porto Alegre. Destaque especial dado à bacia do arroio Dilúvio, onde estudos detalhados de uso e aptidão dos solos cruzados com atributos da paisagem, permitirão um zoneamento ambiental detalhado a nível de unidades de paisagem (entendidas como biótopos), possibilitando um adequado gerenciamento para um desenvolvimento sustentável. Assim, o objetivo geral desse trabalho é realizar estudos de zoneamento ambiental no município de Porto Alegre, visando contribuir ao manejo adequado de áreas, destacando-se o potencial natural destas, através da elaboração de mapas de zoneamento ambiental, constituídos a partir da interpretação de imagens de satélite, servindo este produto como base a delimitações de questões específicas relativas a flora, fauna e geomorfologia da bacia do arroio Dilúvio, na tentativa de definição dos componentes desta.
MATERIAIS E MÉTODOS Áreas de estudo A imagem preliminar de Porto Alegre de uso e ocupação do solo (figura 1) foi obtida a partir da interpretação não supervisionada de uma imagem Spot 1998. Para a escolha das áreas a serem amostradas nos levantamentos foram utilizados os seguintes critérios: a priori, pela cota altimétrica, onde estipularam-se três faixas de altitudes (0-100m, 100-200m e 200-300m) e a rede hidrográfica de contribuintes da bacia do Dilúvio e, a posteriori, para levantamento da flora e da fauna, pela identificação a campo, permitindo a escolha de manchas florestais mais próximas da situação natural. As seguintes áreas foram escolhidas: a) Parque Saint-Hilaire: duas transecções, uma na floresta aluvial de exposição norte na cota de 0-100m e outra na floresta de fundo de vale na cota de 0-100m. b) Morro da Companhia: duas transecções, uma na floresta de terras baixas de exposição sul na cota de 0-100m e outra na floresta de encosta de exposição sul na cota de 100-200m. c) Morro da Polícia: duas transecções, uma na floresta de encosta de exposição sul na cota de 100-200m e outra na floresta de encosta de exposição sul na cota de 200-300m. d) Morro Santana: duas cotas, uma de 100-200m e uma maior que 300m, onde utilizamos os dados de Mohr e Porto (1998). Levantamento da vegetação O levantamento da vegetação seguiu bases fitossociológicas, utilizando-se o método da projeção ortogonal adaptada do quadrante centrado em um ponto (Landau & Porto, 1994), a partir de uma transecção de 30m com orientação norte-sul. Os pontos foram alinhados de dois em dois metros, onde foram amostrados dois indivíduos por ponto. As espécies consideradas foram aquelas que tiveram diâmetro à altura do peito (DAP) maior ou igual a 0,05m. Para cada local forma realizados os levantamentos em duas transeccionais. Levantamento da avifauna A observação da avifauna ocorreu na primavera e verão (setembro à janeiro) através de identificação visual, auxiliados por um binóculo prismático Burshell 10X50 mm, e identificação auditiva (zoofonia). O levantamento das espécies seguiu metodologia ad libitum aconselhável para pequenas áreas, onde o esforço do observador é suficientemente grande para amostrar todas as espécies que utilizam a área estudada, pois o observador passou horas suficientes para amostrar toda a área. As áreas escolhidas foram dois ambientes florestais situados na áreas da bacia do Dilúvio (uma no Parque Saint-Hilaire e outra no Morro Santana). As aves registradas neste trabalho foram identificadas segundo as descrições morfológicas de Sick (1997) e Belton & Dunning (1993). A classificação e a ordem taxonômica das aves, das listas, são de acordo com Belton (1996). Definição dos biótopos Para a definição dos biótopos foram utilizados mapas topográficos do Ministério do Exército na escala 1:50.000 e imagens Landsat TM de 1996. Na delimitação dos biótopos naturais existentes na bacia do arroio Dilúvio, foram utilizadas as seguintes variáveis: morfologia da paisagem, definida pela altitude e declividade; tipo de ocupação e uso do solo; vegetação dominante (tipo de comunidade) e características da avifauna dominante. O cruzamento das informações foi obtido com o auxílio de interpretação da imagem Landsat TM 1996 e por máxima verossimilhança gaussiana (rotina Max Like) do software Idrisi32 e, baseando-se nos estudos fitossociológicos realizados nas unidades que serviram para a análise supervisionada executada na imagem. Na elaboração final do mapa de biótopos naturais da bacia do arroio Dilúvio utilizou-se os softwares Carta Linx versão 1.2 e Corel Draw versão 9.0.
RESULTADOS A figura 1 representa o macrozoneamento ambiental do Município de Porto Alegre onde visualiza-se classes de uso e ocupação do solo. Esta imagem serviu para estabelecer os limites da área a ser estudada (bacia do arroio Dilúvio) e os macrocritérios necessários na escolha das áreas amostrais e no delineamento dos estudos da vegetação. Figura 1 - Classificação prévia do uso do solo por imagem Spot de 1998. |

|
A tabela 1 é resultante dos levantamentos fitossociológicos executados em 8 transeccionais e a relação destes estudos para com os biótopos definidos na figura 2. Os valores comparados são os calculados através do IVI (Índice de Valor de Importância) das espécies vegetais arbóreas por amostra definidos para a Bacia do Arroio Dilúvio Tabela 1 – Índice de Valor de Importância (IVI) e respectivos biótopos de ocorrências das espécies amostradas na Bacia do Arroio Dilúvio, Porto Alegre – RS. Em negrito, destacam-se as espécies que obtiveram os mais altos IVIs. |
|
Espécie |
Biótopo 5 |
Biótopo 6 |
Biótopo 7 |
Biótopo 8 |
|
Aioea saligna |
15,64 |
|||
|
Alchornea triplinervia |
9,15 |
|||
|
Allophylus edulis |
34,65 |
19,40 |
9,0 |
|
|
Banara parviflora |
6,14 |
7,42 |
8,71 |
10,07 |
|
Cabralea cangerana |
31,95 |
|||
|
Campomanesia rombia |
4,06 |
|||
|
Campomanesia xanthocarpa |
14,86 |
|||
|
Casearia decandra |
7,14 |
9,35 |
||
|
Casearia sylvestris |
27,94 |
17,75 |
15,59 |
|
|
Coussapoa microcarpa |
27,22 |
|||
|
Crysophyllum gonocarpum |
27,20 |
|||
|
Crysophylum marginatum |
7,88 |
|||
|
Diospyros inconstans |
18,0 |
20,87 |
||
|
Endlicheria sp. |
3,86 |
|||
|
Enterolobium contortisiliquum |
8,81 |
|||
|
Erythroxylum argentinum |
18,79 |
|||
|
Eugenia rostrifolia |
6,77 |
7,02 |
||
|
Eugenia scheuchiana |
3,59 |
13,97 |
||
|
Faramea marginata |
3,59 |
7,27 |
13,72 |
|
|
Ficus organensis |
27,39 |
|||
|
Gimnanthes concolor |
18,25 |
|||
|
Gomidesia sp. |
15,50 |
|||
|
Guapira opposita |
183,01 |
12,04 |
||
|
Hennecartia omphalandra |
7,34 |
|||
|
Ilex brevicuspis |
10,98 |
|||
|
Inga sessilis |
12,84 |
|||
|
Lithraea brasiliensis |
14,55 |
10,96 |
||
|
Luehea divaricata |
30,98 |
19,34 |
||
|
Machaerium paraguariense |
22,91 |
|||
|
Machaerium stipitatum |
6,87 |
23,20 |
||
|
Matayba elaegnoides |
7,64 |
12,85 |
||
|
Myrcianthes giganthea |
8,56 |
|||
|
Myrcianthes pungens |
7,92 |
|||
|
Myrciaria cuspidata |
7,02 |
81,36 |
||
|
N.I. (Myrtaceae) |
9,45 |
|||
|
Myrsine umbellata |
60,18 |
13,39 |
||
|
Nectandra megapotamica |
6,95 |
|||
|
Nefelia sp. |
3,75 |
|||
|
Ocotea catharinensis |
9,85 |
14,67 |
16,52 |
|
|
Ocotea puberulla |
8,34 |
|||
|
Prunus sellowii |
9,42 |
|||
|
Roupala brasiliensis |
52,80 |
|||
|
Sebastiania brasiliensis |
7,61 |
|||
|
Sebastiania commersoniana |
14,10 |
71,90 |
29,82 |
|
|
Sebastiania serrata |
7,04 |
|||
|
Sorocea bonplandii |
37,72 |
13,55 |
||
|
Symplocos celastrinea |
8,29 |
|||
|
Trichillia clausenii |
42,62 |
15,36 |
||
|
Vitex megapotamica |
19,55 |
6,71 |
||
|
Xylosma sp. |
||||
|
Zanthoxylum rhoifolium |
7,05 |
|
Os valores destacados em negrito representam as espécies dominantes devido ao seu maior valor de IVI, as quais denominam as comunidades vegetais arbóreas nos biótopos 5, 6, 7 e 8 (figura 2). Os demais biótopos não estão contemplados nesta tabela pois representam ambientes não florestais, ambientes construídos ou ambientes aquáticos. Ao total foram identificados 8 biótopos naturais existentes na bacia do arroio Dilúvio: campos baixos e úmidos (Biótopo 1), savanas com butiás e cactáceas (Biótopo 2), campos secos (Biótopo 3), florestas de terras baixas de exposição Sul (Biótopo 4), florestas altas de encostas e de vales em cotas até 100m e exposição Sul (Biótopo 5), florestas baixas de encosta e exposição Sul (Biótopo 6), florestas de terras baixas e exposição Norte (Biótopo 7) e florestas baixas de encosta e exposição Norte (Biótopo 8). Além desses, foram identificados mais dois biótopos referentes a corpos d'água (aquáticos) e os biótopos das áreas construídas (urbanos) que serão analisados na próxima fase deste estudo. A nomenclatura utilizada para a definição dos biótopos, segundo aspectos florísticos e fisionômicos, é proposta por Porto (1998) para o município de Porto Alegre. As espécies dominantes foram utilizadas para denominar as comunidades vegetais dominantes, que serviram de base para identificar os biótopos naturais, sendo as seguintes: - Comunidade de Sebastiania commersionana e Machaerium stipitatum (biótopo 7) - Comunidade de Trichilia clausseni e Sorocea bonplandii (biótopo 5) - Comunidade de Myrciaria cuspidata e Sebastiania commersionana (biótopo 8) - Comunidade de Myrsine umbellata e Guapira opposita (biótopo 6) - Comunidade de Guapira opposita e Myrsine umbellata (biótopo 6) - Comunidade de Guapira opposita e Faramea marginata (biótopo 5) A figura 2, obtida pela interpretação de uma imagem Landsat TM , representa os resultados obtidos, até o momento, para uma visualização das unidades de paisagem (biótopos) interpretadas segundo conceito de Bedê et al. (1994). Nesta figura estão representados nove biótopos naturais, oito dos quais caracterizados segundo critérios de morfologia da paisagem, vegetação e fauna. Nesta figura destacam-se os biótopos urbanos (construídos) que serão objeto de detalhamento em futuras trabalhos. Figura 2 - Mapa preliminar dos biótopos naturais - Bacia do Arroio Dilúvio |
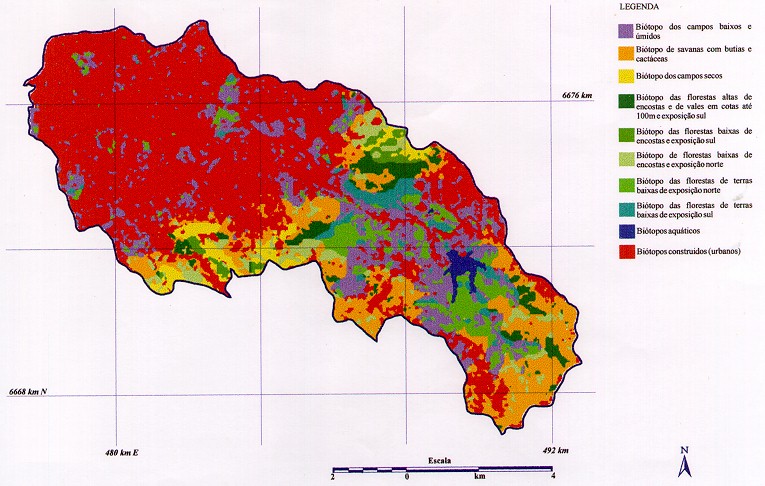
|
A tabela 2, refere-se ao levantamento da avifauna nos biótopos 5 e 7. O biótopo 7 apresentou 65 espécies e o biótopo 5, com uma menor riqueza, apresentou 54 espécies. Tabela 2 - Lista de espécies encontradas nos biótopos 5 e 7. |
|
nome popular |
Nome científico |
Família |
biótopo 5 |
biótopo 7 |
|
|
1 |
Inambuguaçu |
Crypturellus obsoletus |
Tinamidae |
X |
X |
|
2 |
Carrapateiro |
Mivalgo chimachima |
Falconidae |
X |
X |
|
3 |
Araquã |
Ortalis motmot (guttata) |
Cracidae |
X |
X |
|
4 |
Saracura-do-brejo |
Aramides saracura |
Raliidae |
X |
X |
|
5 |
Juriti-pupu |
Leptotila verrauxi |
Columbidae |
X |
X |
|
6 |
Juriti-gemedeira |
Leptotila rufaxilla |
Columbidae |
X |
|
|
7 |
Rolinha-roxa |
Columbina talpacoti |
Columbidae |
X |
|
|
8 |
Alma-de-gato |
Piaya cayana |
Cuculidae |
X |
X |
|
9 |
Anu-preto |
Crotophaga ani |
Cuculidae |
X |
|
|
10 |
Beija-flor-de-topete |
Stephanoxis lalandi |
Trochilidae |
X |
X |
|
11 |
Pica-pau-dourado |
Piculus aurulentus |
Picidae |
X |
X |
|
12 |
Picapauzinho-verde-carijó |
Verniliornis spilogaster |
Picidae |
X |
|
|
13 |
João-velho |
Celeus flavescens |
Picidae |
X |
X |
|
14 |
Pica-pau-do-campo |
Colaptes campestris |
Picidae |
X |
|
|
15 |
Pica-pau-verde-barrado |
Colaptes melanochlorus |
Picidae |
X |
X |
|
16 |
Arapaçu-escamoso |
Lepidocolaptes squamatus |
Dendrocolaptidae |
X |
X |
|
17 |
Arapaçu-verde |
Sittasomus griseicapillus |
Dendrocolaptidae |
X |
X |
|
18 |
Arapaçu-grande |
Dendrocolaptes platyrostris |
Dendrocolaptidae |
X |
X |
|
19 |
João-porca |
Lochmias nematura |
Furnariidae |
X |
X |
|
20 |
Trepador-quiete |
Syndactyla rufosuperciliata |
Furnariidae |
X |
X |
|
21 |
João-de-barro |
Furnarius rufus |
Furnariidae |
X |
|
|
22 |
Arredio-oliváceo |
Cranioleuca obsoleta |
Furnariidae |
X |
X |
|
23 |
João-teneném |
Synallaxis spixi |
Furnariidae |
X |
|
|
24 |
Vira-folhas |
Sclerurus scansor |
Furnariidae |
X |
X |
|
25 |
Pi-puí |
Synallaxis cinerascens |
Furnariidae |
X |
X |
|
26 |
Brujarara-assobiador |
Mackenziaena leachii |
Formicariidae |
X |
X |
|
27 |
Choca-da-mata |
Thamnophilus caerulescens |
Formicariidae |
X |
X |
|
28 |
Tovaca-campainha |
Chamaeza campanisona |
Formicariidae |
X |
|
|
29 |
Chupa-dente |
Conopophaga lineata |
Formicariidae |
X |
X |
|
30 |
Choca-boné-vermelho |
Thamnophilus ruficapilus |
Formicariidae |
X |
|
|
31 |
Bem-te-vi |
Pitangus sulphuratus |
Tyrannidae |
X |
X |
|
32 |
Enferrujado |
Lathrotriccus euleri |
Tyrannidae |
X |
X |
|
33 |
Borboletinha-do-mato |
Phylloscartes ventralis |
Tyrannidae |
X |
X |
|
34 |
Risadinha |
Camptostoma obsoletum |
Tyrannidae |
X |
X |
|
35 |
Guaracava-de-bico-curto |
Elaenia parvirostris |
Tyrannidae |
X |
X |
|
36 |
Tororó |
Todirostrum plumbeiceps |
Tyrannidae |
X |
X |
|
37 |
Bico-chato-orelha-preta |
Tolmomyias sulphurescens |
Tyrannidae |
X |
X |
|
38 |
Patinho |
Platyrinchus mystaceus |
Tyrannidae |
X |
|
|
39 |
Bem-te-vi-rajado |
Myodinastes maculatus |
Tyrannidae |
X |
|
|
40 |
Caneleirinho-preto |
Pachyramphus polychopterus |
Tyrannidae |
X |
X |
|
41 |
Irrê |
Myarchus swainsoni |
Tyrannidae |
X |
X |
|
42 |
Tuque |
Elaenia mesoleuca |
Tyrannidae |
X |
|
|
43 |
Caneleirinho-verde |
Pachyramphus viridis |
Tyrannidae |
X |
X |
|
44 |
Suiriri |
Tyrannus melancholicus |
Tyrannidae |
X |
|
|
45 |
Dançador |
Chiroxiphia caudata |
Pipridae |
X |
X |
|
46 |
Corruíra |
Troglodytes aedon |
Troglodytidae |
X |
X |
|
47 |
Sabiá-coleira |
Turdus albicollis |
Muscicapidae/Turdinae |
X |
X |
|
48 |
Sabiá-laranjeira |
Turdus rufiventris |
Muscicapidae/Turdinae |
X |
X |
|
49 |
Sabiá-poca |
Turdus amaurochalinus |
Muscicapidae/Turdinae |
X |
X |
|
50 |
Sabiá-una |
Platycichla flavipes |
Muscicapidae/Turdinae |
X |
|
|
51 |
Pitiguari |
Cyclarhis gujanensis |
Vireonidae/Cyclarhinae |
X |
X |
|
52 |
Juruviara |
Vireo olivaceus |
Vireonidae/Vireoninae |
X |
X |
|
53 |
Tico-tico |
Zonotrichia capensis |
Emberezidae/Emberezinae |
X |
X |
|
54 |
Quete |
Poospiza lateralis |
Emberezidae/Emberezinae |
X |
X |
|
55 |
Trinca-ferro-verdadeiro |
Saltator similis |
Emberezidae/Cardinalinae |
X |
|
|
56 |
Saíra-preciosa |
Tangara preciosa/peruviana |
Emberezidae/Thraupinae |
X |
X |
|
57 |
Gaturamo-rei |
Euphonia musica |
Emberezidae/Thraupinae |
X |
X |
|
58 |
Fim-fim |
Euphonia chlorotica |
Emberezidae/Thraupinae |
X |
X |
|
59 |
Saíra-viúva |
Pipraeidea melanonota |
Emberezidae/Thraupinae |
X |
X |
|
60 |
Gaturamo-serrador |
Euphonia pectoralis |
Emberezidae/Thraupinae |
X |
X |
|
61 |
Sanhaçu-cinzento |
Thraupis sayaca |
Emberezidae/Thraupinae |
X |
|
|
62 |
Tiê-preto |
Tachyphonus coronatus |
Emberezidae/Thraupinae |
X |
|
|
63 |
Sanhaçu-frade |
Stephanophorus diadematus |
Emberezidae/Thraupinae |
X |
|
|
64 |
Cais-cais |
Euphonia chalybea |
Emberezidae/Thraupinae |
X |
|
|
65 |
Pula-pula-assobiador |
Basileuterus leucoblepharus |
Emberezidae/Parulinae |
X |
X |
|
66 |
Mariquita |
Parula pitiayumi |
Emberezidae/Parulinae |
X |
X |
|
67 |
Pula-pula |
Basileuterus culicivorus |
Emberezidae/Parulinae |
X |
X |
|
68 |
Pia-cobra |
Geothlypis aequinoctialis |
Emberezidae/Parulinae |
X |
|
|
69 |
Cambacica |
Coereba flaveola |
Emberizidae/Coerebinae |
X |
X |
|
DISCUSSÃO A utilização de critérios morfológicos da paisagem parece ter sido a base inicial adequada para definição das unidades de paisagem, possibilitando desta forma, uma macrovisão de situações que favorecem o estabelecimento de recursos naturais distintos de flora, fauna e substrato. Segundo Bedê et al. (1994) "O mapeamento dos biótopos nada mais é que o registro cartográfico de parcelas de uma paisagem sob um enfoque abrangente, integrando aspectos do meio físico, biológico e antrópico na forma de um mosaico de tipologias (tipos de biótopos), que caracteriza uma superfície do ponto de vista ambiental, como um todo." A análise da vegetação revela , através de estudos fitossociológicos, a existência de comunidades estruturadas e de composição florística esperada para as áreas naturais de Porto Alegre. Porto (1998) define 8 classes de vegetação nativa para o município de Porto Alegre, as quais são claramente identificadas em interpretação de uma imagem Landsat TM , visualizando-se no presente trabalho todas as espécies que foram detectadas como dominantes nas diferentes comunidades constatadas no trabalho acima referido. A composição florística do estrato das árvores deixa transparecer a sua originalidade como vegetação, no entanto, em muitos casos, o estrato herbáceo e o arbustivo estão alterados, tendo em vista, que os remanescentes da vegetação encontram-se muito próximos da urbanização ou à áreas de utilização agropecuária. A definição preliminar dos biótopos naturais nos revela, espacialmente, uma situação cuja vegetação tem seu papel importante na retenção do substrato e na disponibilidade de alimento e abrigo para a fauna, podendo estas unidades serem as informações ideais para o manejo e conservação dos recursos naturais em um ambiente urbanizado. Atualmente, os estudos em estrutura da paisagem e, principalmente, em fragmentação de habitats, vêem utilizando elementos da fauna como objeto de investigação. Como exemplo desta tendência, pode-se citar Flather & Sauer (1996) que encontrou diferentes comportamentos de tolerância à alteração da paisagem em grupos diferentes de pássaros migratórios em uma larga escala. Em outro estudo, Villard & Taylor (1994) verificou que pássaros tolerantes à fragmentação de habitat detectam e colonizam novos habitats mais rápido que pássaros intolerantes. Desta forma, observa-se que as aves, por serem organismos muito móveis e ativos, respondem diretamente às características estruturais da paisagem. Uma análise preliminar sugere que a maior riqueza encontrada no biótopo 7 está associada à diversidade paisagística deste, pois as bordas deste biótopo estão em contato com campo e banhado; esta situação não ocorre no biótopo 5. Desta forma, os dados de riqueza das aves , revelam diferenças entre biótopos, como é o caso do biótopos 7 e 5, deixando evidenciar, ao mesmo tempo, informações sobre a influência da matriz da paisagem e suas áreas da contato entre biótopos, podendo servir como indicadores para corredores em um planejamento integrado de paisagem. O geoprocessamento realizado com os dados obtidos possibilitou a integração e a visão sintética necessária, modelando, desta forma, uma situação que mesmo preliminar, informa sobre unidades da paisagem que têm características ambientais homogêneas e que apresentam funcionalidades próprias, as quais devem ser respeitadas no planejamento de espaços. Tais considerações possibilitam aos tomadores de decisão chegarem próximo ao desejável, isto é, a um planejamento ecologicamente sustentado.
REFERÊNCIAS BIBLIOGRÁFICAS BEDÊ, L.C.; WEBER, N.; RESENDE, S.; PIPER, W.; SCHULTE, W. 1994. Manual para o mapeamento de biótopos no Brasil. Belo Horizonte: Ed. Brandt Meio Ambiente Ltda. 100 p. il. Belton, W. 1996. Aves do Rio Grande do Sul: Distribuição e Ecologia. UNISINOS. São Leopoldo. Belton, W. e Dunning, J. S. 1993. Aves silvestres do Rio Grande do Sul. Fundação Zoobotânica do RS, Porto Alegre, RS. BRACK, P., RODRIGUES, R. S., SOBRAL, M., LEITE, S. L. de C. 1998. Árvores e arbustos na vegetação natural de Porto Alegre, no Rio Grande do Sul, Brasil. Iheringia, vol II, no. 51, p. 137-166. FLATHER C. H., SAUER J. R. Using landscape ecology to test hypotheses about large-scale abundance patterns in migratory birds. Ecology, vol 77(1), p. 28-35. 1996. LANDAU, E. C; PORTO, M.L. Ecologia de paisagem da região do vale do Rio dos Sinos – Santa Cristina do Pinhal, Parobé/RS em bases fitossociológicas e de geoprocessamento. Porto Alegre: UFRGS, 1994. Dissertação (Mestrado em Ecologia), Instituto de Biociências, Curso de Pós-Graduação em Ecologia. MOHR, F. V. e PORTO, M. L. 1998. Cap. 8.1.: Morro Santana: o verde luxuriante nas ncostas íngremes. In: MENEGAT; R., PORTO, M. L.; CARRARO, C. C.; FERNANDES, L. A. D. 1998. Atlas Ambiental de Porto Alegre. Porto Alegre: Ed. da UFRGS. P. 81. PORTO, M. L. 1998. Cap. 5: As formações vegetais: evolução e dinâmica da conquista. In: MENEGAT; R., PORTO, M. L.; CARRARO, C. C.; FERNANDES, L. A. D. 1998. Atlas Ambiental de Porto Alegre. Porto Alegre: Ed. da UFRGS. P. 47. PORTO, M. L e OLIVEIRA, M. L. A. 1998. Cap. 14.5.: O impacto ambiental no Arroio Dilúvio: a enfermidade da água. In: MENEGAT; R., PORTO, M. L.; CARRARO, C. C.; FERNANDES, L. A. D. 1998. Atlas Ambiental de Porto Alegre. Porto Alegre: Ed. da UFRGS. P. 47. Sick, H. 1997. Ornitologia Brasileira. Editora Nova Fronteiras. Rio de Janeiro. VILLARD M. A, TAYLOR P. D. Tolerance to habitat fragmentation influences the colonization of new Habitat by forest birds. Oecologia, vol. 98(3-4), p. 393-401. 1994.
|
